|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Diaptomoidea ( Superfamille ) |
|
|
|
Pontellidae ( Famille ) |
|
|
|
Anomalocera ( Genre ) |
|
|
| |
Anomalocera patersoni Templeton, 1837 (F,M) | |
| | | | | | | Syn.: | Irenaeus Patersonii : Claus, 1863 (p.206);
Pontia Pattersonii : Kröyer, 1849 (in Damkaer & Damkaer, 1979, p.41);
no Anomalocera patersoni : Shih & al., 1971 (p.43) | | | | Ref.: | | | Lubbock, 1853 b (p.159, 164, fig.M); Brady, 1878 (p.75, figs.F,M); Bourne, 1889 (p.149); Giesbrecht, 1892 (p.479, figs.F,M); Giesbrecht & Schmeil, 1898 (p.145); Wheeler, 1901 (p.181, figs.F,M); Sars, 1902 (1903) (p.139, figs.F,M); Farran, 1908 b (p.87); Lysholm, 1913 (p.7); Pesta, 1920 (p.544); Sars, 1925 (p.352); Farran, 1926 (p.292); Rose, 1929 (p.44); Sewell, 1932 (p.350, Rem.); Wilson, 1932 a (p.142, figs.F,M); Rose, 1933 a (p.257, figs.F,M); Jespersen, 1934 (p.119); 1940 (p.67, fig.7); Wilson, 1942 a (p.171, fig.F); Lysholm & al., 1945 (p.42); Porumb, 1970 (fiche CIESMM n°35); Razouls, 1972 (p.95, Annexe: p.90); Silas & Pillai, 1973 (p.780, Rem.); Schnack, 1989 (p.137, tab.1, fig.6: Md); Sazhina, 1985 (p.76, figs.N); Huys & Boxshall, 1991 (p.66, fig.); Ianora & Santella, 1991 (p.387, diapause eggs, figs.); Ianora & al., 1992 (p.401, fig., Rem.); Barthélémy & al., 1998 (p.721, genital area); Barthélémy, 1999 a (p.9, 75, figs.7, 37, B-C: glande coquillière); Bradford-Grieve, 1999 b (p.181, Déf.); Boxshall & Halsey, 2004 (p.163, figs.F,M); Avancini & al., 2006 (p.103, Pl. 72, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.488, figs.F,M, Rem.); Laakmann & al., 2013 (p.862, figs.1, 2, 3, 5, Table 1, 2, 3, mol. Biol.) | 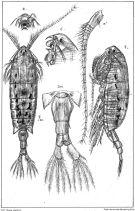 issued from : G.O. Sars in An Account of the Crustacea of Norway. Vol. IV. Copepoda Calanoida. Published by the Bergen Museum, 1903. [Pl. XCII]. Female.
|
 issued from : G.O. Sars in An Account of the Crustacea of Norway. Vol. IV. Copepoda Calanoida. Published by the Bergen Museum, 1903. [Pl.XCIII]. Female. Nota: M = Md; m = Mx1; mp1 = Mx2; mp2 = Mxp
|
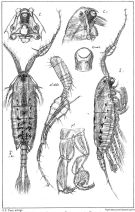 issued from : G.O. Sars in An Account of the Crustacea of Norway. Vol. IV. Copepoda Calanoida. Published by the Bergen Museum, 1903. [Pl.XCIV]. Male. Nota: C = forehead (lateral right side); O.ventr = ventral eye.
|
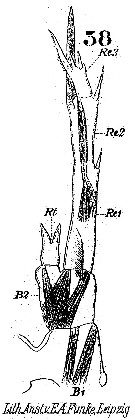 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Fig.38]. Female: 38, P5 (posterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Fig.37]. Male: P5 (posterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Fig.10]. Female: 10, Md (posterior view). Re = exopod.
|
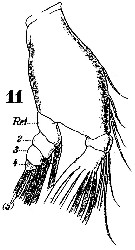 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Fig.11]. Female: 11, Md (mandibular palp, anterior view). Re = exopod.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Fig.17]. Female: 17, Md (masticatory edge). Si = inner seta.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Fig.19]. Female: 19, Mxp (anterior view). B = basipod; L = lobes; Ri = endopod; Si = inner seta; Se = outer seta.
|
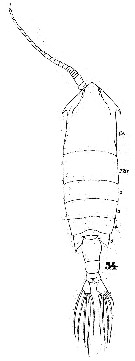 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.40, Fig.54]. Female: 54, habitus (dorsal).
|
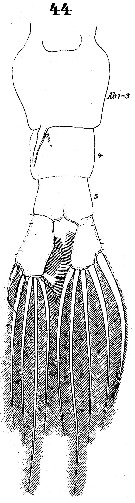 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.40, Fig.44]. Female: 44, urosome (ventral).
|
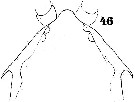 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.40, Fig.46]. Female: 46, forehead (dorsal).
|
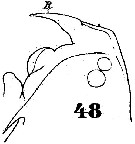 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.40, Fig.48]. Female: 48, forehead (lateral). R = rostrum.
|
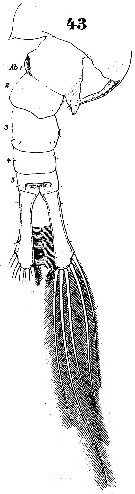 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.40, Fig.43]. Male: 43, Th5 and urosome (dorsal). Ab 1 = urosomal somite 1.
|
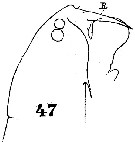 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.40, Fig.47]. Male: 47, forehead (lateral).
|
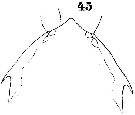 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.40, Fig.45]. Male: 45, forehead (dorsal).
|
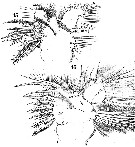 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Figs.15, 16]. Female: 15, Mx1 (posterior view); 16, idem (anterior view). Le = outer lobe; Li = inner lobe; Ri = endopod; Re = exopod; B2 = basipodal segment 2.
|
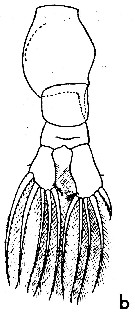 issued from : W.M. Pennell in Can. J. Zool., 1976, 54. [p.1665, Fig.2b]. Female (from G. of St Lawrence): b, urosome (dorsal).
|
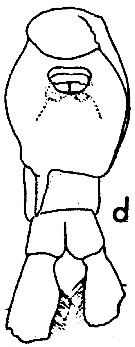 issued from : W.M. Pennell in Can. J. Zool., 1976, 54. [p.1667, Fig.5d]. Female: d, urosome (ventral).
|
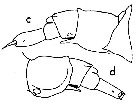 issued from : W.M. Pennell in Can. J. Zool., 1976, 54. [p.1666, Fig.3 c, d]. Female: c, urosome (lateral right side); d, idem (lateral left side).
|
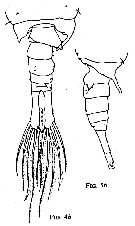 issued from : W.M. Pennell in Can. J. Zool., 1976, 54. [p.1667, Fig.4b, Fig.5b]. Male: 4b, urosome (dorsal); 5b, idem (lateralright side).
|
 issued from : W.M. Pennell in Can. J. Zool., 1976, 54. [p.1668, Fig.6]. Regression of length of first prosome segment on length of genital segment in A. opalus and females.
|
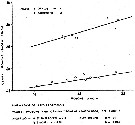 issued from : W.M. Pennell in Can. J. Zool., 1976, 54. [p.1668, Fig.6]. Regression of length of first prosome segment on length of genital segment in A. opalus and males.
|
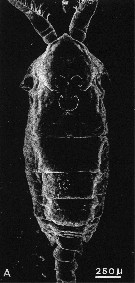 issued from : A. Ianora, A. Miralto & S. Vanucci in Mar Biol., 1992, 113. [p.402, Fig.1, A]. SEM micrographs. Male: A, surface attachment structure (mass of fine setules arranged in three semicircles on a flattened area of the anterodorsal surface of the cephalosome). Nota: The structure is a complex, double horsehoeshaped formation in both copepodites and adults, the two semicircles are oriented longitudinally, with the concave surface facing each other. The general shape of this forlmation is similar in both males and females. Both semicircles consist of a thick band of setules or twisted hairs with a unramous base and branched apex with blunt ends. large pore and peg sensilla are often situated close to the structure. Not all pontellids posses this structure.. The structure is continuous with the cuticle and has no underlying sub-integumental stuctures. The main shaft of setules or hairs which emerge dorsally from the integument consists of an outer layer (epicuticule) and an inner layer (upper procuticle). The structure may be considered one of a variety of cuticular structures described for the integument of Crustacea, including the velcro-like, cincinnuli adhesions observed on some pleopod appendages of decapods and euphausiids and which apparently are involved in swimming or the hooking-type pereiopods in commensal decapod crustaceans for holding onto the host's body (Fujino, 1975) These adhesive structures are not associated with neurosecretory celles; their functions are determined by the precise shape and mechanical properties of the arthropod cuticle (Bouchon & Chaigneau, 1991) This structure can be considered a remarkable adaptation to life on the surface. This structure presumably facilitates attachment to the surface by increasing the surface area of adhesion. According to Pennell (1973), in order to attach itself to the surface film Anomalocera opalus appears to bump upward against the film with its dorsal surface . Once the structure breaks the surface it becomes dry and apparently water-repellent
|
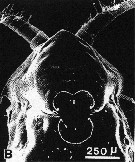 issued from : A. Ianora, A. Miralto & S. Vanucci in Mar Biol., 1992, 113. [p.402, Fig.1, B]. SEM micrographs. Femle: B, surface attachment structure.
|
 issued from : A. Ianora, A. Miralto & S. Vanucci in Mar Biol., 1992, 113. [p.402, Fig.1, E]. SEM micrographs. E, central mass of filaments comprising the .surface attachment structure
|
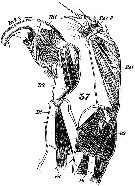
 issued from : M. Rose in Result. Camp. Scient. Price Albert Ier, Monaco, 1929, 78. [Pl.III, Figs.7]. Male: D', habitus (dorsal); L, idem (lateral); T, forehead (dorsal); R', idem (lateral); Th5, process of the last right thoracic segment; Ab', urosome (lateral); P5 d, right P5; P5 g, left P5.
|
 issued from : R. Huys & G.A. Boxshall in Copepod Evolution. The Ray Society, 1991, 159. [p.66, Fig.2.2.13, B]. From English Channel: Plymouth: B, exopod of A2.
|
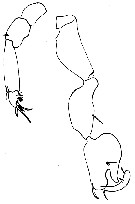 issued from : C; Razouls, unpublished (pers. coll.), in Toulon harbour, 31 october 1964. Male: P5.
|
 issued from : S.B. Schnack in Crustacean Issue, 1989, 6. [p.143, Fig.6: 12]. 12, Anomalocera patersoni (from off NW Africa, upwelling region): Cutting edge of Md.
|
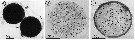 issued from : A. Ianora & L. Santella in Mar. Biol., 1991, 108. [p.388, Fig.1, A-C]. Anomalocera patersoni from the Gulf of Naples (Italy). A: Diapause eggs enveloped by transparent pellicle. B: Histological section of subitaneous egg with thin outer coat and C of diapause egg with multi-layer coat.
|
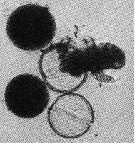 issued from : A. Ianora & L. Santella in Mar. Biol., 1991, 108. [p.392, Fig.5]. Anomalocera patersoni from the Gulf of Naples (Italy). Hatching of diapause eggs after 7 monthes of dormancy. Nauplius visible.
|
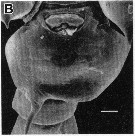 issued from : R.-M. Barthélémy in Thèse Doct. Univ. Provence (Aix-Marseille I), 1999. [Fig.7, B ]. Female (from Gulf of Marseille, NW Mediterranean Sea): external ventral view genital double-somite. Scale bar: 0.050 mm.
|
 issued from : R.-M. Barthélémy, C. Cuoc, D. Defaye, M. Brunet & J. Mazza in Phil. Trans. R. Soc. Lond., B, 1998, 353. [p.733, Fig.63]. Schematic representation of external genital area in the species studied; Dashed line, limit of the anterior pad and lateral thickenings; shading, posterior pad; dots, genital operculum.
| | | | | Ref. compl.: | | | Mrazek, 1902 (p.510); Pearson, 1906 (p.30); Sars, 1909 b (p.17); Chatton, 1920 (p.17); Rose, 1925 (p.152); Wilson, 1932 (p.21); Massuti Alzamora, 1942 (p.96, Rem.); Sewell, 1948 (p.506, 522); C.B. Wilson, 1950 (p.165); Fleury, 1950 (p.47, fig.2); Østvedt, 1955 (p.15: Table 3, p.76); Demir, 1959 (p.176, Rem.); Lacroix, 1960 (p.11, 25); Deevey, 1960 (p.5, Table II, annual abundance); Wickstead, 1962 (p.546, food & feeding); Grice & Hart, 1962 (p.287, 295: Rem.); Lacroix & Bergeron, 1963 (p.59, Tableau III, IV); Duran, 1963 (p.24); Giron-Reguer, 1963 (p.53); Rice, 1964 (p.163, hydrostatic pressure effects); Mazza, 1966 (p.72); 1967 (p.328, 329); Ehrhardt, 1967 (p.741, geographic distribution, Rem.); Evans, 1968 (p.11); Matthews, 1967 (p.159, Table 1, Rem.); Sherman & Schaner, 1968 (p.583, fig.2); Champalbert, 1969 (p.346, vertical microdistribution); 1969 a (p.595); Dowidar & El-Maghraby, 1970 (p.267); Marcus, 1970 (p.10); Kovalev, 1970 a (p.87, Table 1, 2, comparison hyponeustonic & planctonic forms); Champalbert & Gaudy, 1971 (p.71); 1972 (p.159, respiration respiration vs. temperature); Apostolopoulou, 1972 (p.328, 367); Macdonald & al., 1972 (p.213, hydrostatic pressure effect); Bainbridge & Forsyth, 1972 (p.21, Table I: indictor species, Table II: correlation analysis); Lefèvre-Lehoërff, 1972 (p.1681); Champalbert & al., 1973 (p.529, CHN composition); Champalbert, 1973 (p.315, respiration v.s. temperature-weight); 1973 a (p.208, phototropism); Weikert, 1975 (p.137, chart); Fernandez, 1975 (p.1, fig.15, 16, metabolism/lux), Kerambrun & Champalbert, 1975 (p.1019, biochimy); Champalbert & al., 1976 (1978) (p.263, geographic size comparison); Champalbert & Kerambrun, 1978 (p.215, weight-CHN); Fernandez, 1978 (p.97, metabolism/food, Rem.: Table 19); Small & al., 1979 (p.233, fecal pellets); Porumb, 1980 (p.167); Samain & al., 1980 (p.225, polluant effects); 1981 (p.35, Rem.: polluant effect); Vives, 1982 (p.295); Kerambrun & Champalbert, 1982 (p.17, C/N/H & proteins composition); Kovalev & Shmeleva, 1982 (p.85); Moraitou-Apostolopoulou, 1985 (p.303, occurrence/abundance in E Mediterranean Sea, Rem.: p.316); Regner, 1985 (p.11, Rem.: p.38); Williams & Collins, 1985 (p.27); Brenning, 1985 a (p.28, Table 2); 1987 (p.30, Rem.); Gorsky & al., 1988 (p.133, Table I, C-N composition); Poulet & al., 1989 (p.1325, Table 2: vitamin content); Citarella, 1989 (p.123, abundance); Valdes & al., 1990 (tab.2); Fransz & al., 1991 (p.8); Séguin & al., 1993 (p.23); Kouwenberg, 1994 (tab.1); Marcus, 1996 (p.144); Zauke & al., 1996 (p.141, Table 7, metal bioaccumulation); Mauchline, 1998 (p.505, tab.40, 46); Hure & Krsinic, 1998 (p.102); Gilabert & Moreno, 1998 (tab.1, 2); Siokou-Frangou, 1999 (p.478) ; d'Elbée, 2001(tabl. 1); Holmes, 2001 (p.30); Fransz & Gonzalez, 2001 (p.255, tab.1); Sameoto & al., 2002 (p.12); Beaugrand & al., 2002 (p.179, figs.5, 6); Vukanic, 2003 (p.139, tab.1); Kovalev, 2003 (p.47); Zagorodnyaya & al., 2003 (p.52)Bonnet & Frid, 2004 (p.485, fig.5); CPR, 2004 (p.49, fig.135); Valdés & al., 2007 (p.104: tab.1); Cabal & al., 2008 (289, Table 1); Brylinski, 2009 (p.253: Tab.1, p.256: Rem.); Eloire & al., 2010 (p.657, Table II, temporal variability); Dvoretsky & Dvoretsky, 2010 (p.991, Table 2); Mazzocchi & Di Capua, 2010 (p.427); S.C. Marques & al., 2011 (p.59, Table 1); Alvarez-Fernandez & al., 2012 (p.21, Rem.: Table 1); Gubanova & al., 2013 (in press, Rem.: p.7, Table 4); Belmonte & al., 2013 (p.222, Table 2, abundance vs stations); Kürten & al., 2013 (p.167, Table 1, C:N, fatty acid); Mazzocchi & al., 2014 (p.64, Table 4, abundance); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Ben Ltaief & al., 2017 (p.1, Table III, Summer relative abundance); El Arraj & al., 2017 (p.272, table 2); Baumgartner & Tarrant, 2017 (p.387, resting eggs, Rem.); Belmonte, 2018 (p.273, Table I: Italian zones) | | | | NZ: | 9 | | |
|
Carte de distribution de Anomalocera patersoni par zones géographiques
|
| | | | | | | | | | | |  Carte de 1996 Carte de 1996 | |
 issued from : P. Kerambrun & G. Champalbert in J. Exp. Mar. Biol. Ecol., 1982, 63. [p.20, Fig.1]. issued from : P. Kerambrun & G. Champalbert in J. Exp. Mar. Biol. Ecol., 1982, 63. [p.20, Fig.1].
Evolution of C, H, N elementary composition in relation to the dry weight from juvenils (black triangles), males (black squares and females (black circles) in the Gulf of Marseille (France). |
| | | | Loc: | | | Cape Verde Is., off Mauritania, Moraccan coast, off Madeira, Portugal (Mondego estuary), Bay of Biscay, Arcachon, Delaware Bay, Chesapeake Bay, G. of Maine, Woods Hole, Georges Bank, Bay of Fundy, Northumberland Strait, Baie des Chaleurs, G. of St. Lawrence, off Nova Scotia E, Strait of Belle Isle, W Greenland (Sukkertoppen), Iceland, Faroe Is., Mourmansk, W & S Barents Sea, Irish coasts, Bristol Channel, Norway Sea, W Norway, Raunefjorden, NE Scotland, North Sea, German Bight, English Channel, Roscoff, Morlaix estuary, Pas de Calais, Medit. (Alboran Sea, Alger, Castellon, Baleares, off Barcelona, Banyuls, Marseille, Toulon (Harbour), off Monaco, Villefranche-s-Mer, Ligurian Sea, Napoli, Tyrrhenian Sea, Gulf of Taranto, G. of Gabes, Malta, Adriatic Sea, Aegean Sea, Bosphore, Lebanon Basin, Alexandria, Marmara Sea, Black Sea (rare), Mozambique Channel (in Thompson, 1900 c), Indian (Maldives in Wolfenden, 1905), Philippines (in C.B. Wilson, 1950, p.165), Japan (in Wilson, 1942 a), N Japan (in Marukawa, 1921), off E japan, NE Samoa Is. (in Wilson, 1942 a, p.130, 166) | | | | N: | 141 | | | | Lg.: | | | (38) F: 3,18-3,12; (45) F: 3,25-3; M: 3-2,5; (46) F: 4,1-3,5; M: 3,2-3; (65) F: 3,2; M: 3; (188) F: 4,1-3,5; M: 3-3,2; (449) F: 4,1-3,2; M: 4-3; (789) [Gulf of Marseille] F: 3,85 ± 0,03; M: 3,51± 0,03; [Black Sea: Bosphore] F: 3,36 ± 0,02; M: 3,07 ± 0,03; (920) F: 4; M: 3,62; (1110) M: 3,6; {F: 3,00-4,10; M: 2,50-4,00} | | | | Rem.: | épipélagique.
Cette forme est souvent observée en Atlantique nord, et est présente en Mer du Nord par pénétration des eaux atlantiques (espèce indicatrice). Elle semble très rare dans l'Indien et le Pacifique (voir remarque in Sewell, 1948, p.432).
D'après Porumb (1970) cette espèce eurytherme se rencontre dans toutes les régions de la Mer Noire dont il constitue l'un des principaux composants de l'hyponeuston, principalement près des côtes. Parfois en exemplaire isolé jusqu'à 10-15 m de profondeur. La durée de l'incubation des oeufs est de 2-3 jours, les stades naupliens se rencontrent en automne. L'étude de la composition biochimique montre la capacité de cette espèce à concentrer l'argent dans ses tissus.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 25/10/2022 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 14 juin 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
