|
|
 |
Fiche d'espèce de Copépode |
|
|
Cyclopoida ( Ordre ) |
|
|
|
Oithonidae ( Famille ) |
|
|
|
Oithona ( Genre ) |
|
|
| |
Oithona hebes Giesbrecht, 1891 (F,M) | |
| | | | | | | Syn.: | no Oithona hebes : Grandori, 1913; Wilson, 1942 a (p.196); Gonzalez & Bowman, 1965 (part., figs.19, 20a); Wellershaus, 1969 (p.276, figs.F,M); Goswami, 1975; Fonseca & Björnberg, 1975 (1976) (p.130); Zacarias & Zoppi de Roa, 1981 (p.109, figs.N, juv.); Reddy & Radhakrishna, 1984;
? Oithona sapucaiae Oliveira, 1945 (p.464, figs.F,M);
Oithona canhanhae Oliveira, 1945 (p.465);
Oithona oraemaris Oliveira, 1946 (p.478);
Oithona ovalis Herbst, 1955 (p.214, Descr.F, figs.F); Björnberg, 1963 (p.76, Rem.); Wellershaus, 1970; Björnberg, 1972; Santos, 1973; Fanta, 1976; Fonseca & Björnberg, 1976; Medeiros & Björnberg, 1978; Valentin & al., 1978; Fonseca & Prado, 1979 ; Shuvalov, 1980 (p.158, figs.F); Björnberg & al., 1981; Paranagua & al., 1986 (p.359); Ferrari & Dahms, 2007 (p.31, 32, 33, Rem. N); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2);
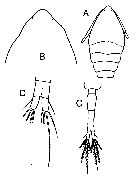
Oithona minuta : Coker & Gonzalez, 1960 | | | | Ref.: | | | Giesbrecht, 1892 (p.538, 549, 774, figs.F); Rosendorn, 1917 a (p.44, Rem. F); Pesta, 1920 (p.551); Kiefer, 1929 g (p.9, Rem.F); Rose, 1933 a (p.280, figs.F); Sewell, 1948 (p.393); Lindberg, 1955 a (p.467: Rem.); Gonzalez & Bowman, 1965 (part., p.269, Rev., fig.20b); Razouls, 1972 (p.95, Annexe: p.109); Björnberg, 1972 (p.87); Bowman, 1975 (Rem.: p.134, figs.F); Ferrari, 1977 (p.406, figs.F,M, Rem.); Shuvalov, 1980 (p.156, figs.F); Ferrari & Bowman, 1980 (p.13, figs.F, M, Rem.); Reid, 1985 a (fig.F, Rem); Rocha, 1986 (p.96, Redescr.F,M, figs. F,M, Rem.); Bradford-Grieve & al., 1999 (p.886, 966, figs.F,M); Vives & Shmeleva, 2010 (p.61, figs.F,M, Rem.) | 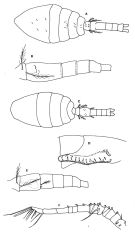 issued from : F.D. Ferrari in Proc. Biol. Soc. Washington, 1977, 90 (2). [p.406, Fig.4]. From Tutuila Island, American Samoa. Female: A, habitus (dorsal); B, urosome (lateral). Male: C, habitus (dorsal); D, cephalosome (ventrolateral); E, urosome (lateral); F, A1.
|
 issued from : F.D. Ferrari in Proc. Biol. Soc. Washington, 1977, 90 (2). [p.409, Fig.5]. From Tutuila Island, American Samoa. Female: A, caudal ramus (ventral); B, Md. Nota: Nota: Endopod of Md with 4 setae, ultimate with setules; basipod 2 with 2 thick spines armed with numerous fine denticles. Male: C, caudal ramus (dorsal); D, Md. Nota: Endopod of Md with 4 setae, distal 2 spines of basipod reduced, only one armed with denticles.
|
 issued from : F.D. Ferrari in Proc. Biol. Soc. Washington, 1977, 90 (2). [p.405, Fig.3]. From Tutuila Island, American Samoa. Female: C, Mx2; D, P4 (anterior). Nota: Mx2 without notch or denticles on large spines. Endopod of P4 with 2 modified setae with membranous flange.
|
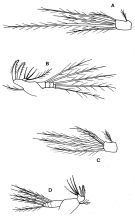
 issued from : F.D. Ferrari & T.E. Bowman in Smiths. Contr. Zool., 1980, 312. [p.13, Fig.7]. Female (from Caribbean area): a, forehead (lateral); b, urosome (lateral right side); c, P4. Nota: Head distinctly quadrate in lateral view. Distal seta of endopodal segment 2 of P4 thicker than proximal of endopodal segment3 (but not so great a difference as shown in figure 3D of Ferrari, 1977). P5 with reduced seta at base. Knob near genital opening with a single well)developed spine (not a small point). Male: posterior portion of cephalosome (lateral right side); e, anterodorsal \"pore signature\" clusters from left sides of 3 other specimens. Nota: P5 with reduced seta at its base.
|
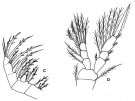
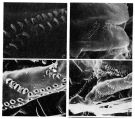 issued from : F.D. Ferrari & T.E. Bowman in Smiths. Contr. Zool., 1980, 312. [p.24, Fig.15 b-e]. Male (SEM micrographs of parts of \"pore signature\"): b, anterodorsal cluster in the center of the field (left side). The integument has cracked along the dorsal horizontal row (to the left) although the first fex organs of that row can be seen. Ventrally the integument has curled and only the top organs of the columns can be seen. c, view of the anterodorsal cluster; each integumental organ consists of a pore, guarded on one side by a thin ridge (the relief of this ridge appears variable for those organs in the cluster. It is this ridge that is visible under the light microscope). Short hairs can be seen associated with most ridges under the light microscope (they are absent in the micrograph). d, the right side of another male (badly distorted due to buckling). Organs of the antero-dorsal cluster are visible in the top right corner; the dorsal horizontal row passes obliquely toward the lower left corner; these pores also seem to have distinct ridges associated with them. In the center are several columns that extend into horizontal series. e, another view of the conjunction of column to horizontal series; distint ridges also appear to be associated with the pores of columns (in contrast, the pores of horizontal series are surrounded by slightly thickened integument). Remarks: Variation in the \"pore signature\" can be noted in specimens examined with the light microscope and we have not had enough specimens to study differences between inter- and intraspecific variation. For this reason we have not been able to find consistent differences between the \"pore signature\" of O. fonsecae and O. hebes.
|
 issued from : T.E. Bowman in Chesapeake Sc., 1975, 16 (1)/ [p.135, Fig1, i-j]. Female (from Gulf of Guayaquil, Ecuador): i, forehead (dorsal); j, idem (lateral). Nota: Md has 5 setae on the endopod and a pair of blunt setae on the 2nd basipod.
|
 issued from : V.S. Shuvalov in Opred. Faune SSSR, Nauka, Leningrad, 1980, 125. [p.159, Fig.46]. As Oithona ovalis. After Herbst, 1955. Female: A, habitus (dorsal); B, A1; C, thoracic segment 5 and genital segment; D, A2; E, Mx1; F, Md; G, Mx2.
|
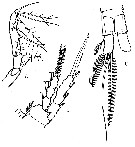 issued from : V.S. Shuvalov in Opred. Faune SSSR, Nauka, Leningrad, 1980, 125. [p.160, Fig.47]. As Oithona ovalis. After Herbst, 1955. Female: A, Mxp; B, P4; C, anal segment and caudal rami.
|
 issued from : V.S. Shuvalov in Opred. Faune SSSR, Nauka, Leningrad, 1980, 125. [p.157, Fig.45]. Female: A, prosome (dorsal); B, forehead (dorsal); C, end of prosame and (dorsal); D, anal segment and caudal rami.
|
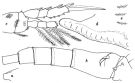
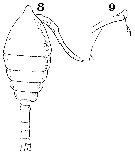 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Figs.8, 9]. Female: 8, habitus (dorsal); 9, foprehead (lateral).
| | | | | Ref. compl.: | | | Carazzi & Grandori, 1912 (p.15, 38); Wilson, 1942 a (p.196); Mazza, 1966 (p.73); Evans, 1968 (p.14); Vaissière & Séguin, 1980 (p.23, tab.1); Vives, 1982 (p.295); Kovalev & Shmeleva, 1982 (p.85); Dussart, 1984 (p.25, 64: occurrence): Dussart & Defaye, 1985 (p.10); Seguin & al., 1993 (p.23); Lopes, 1994 (tab.1); Webber & Roff, 1995 (tab.1); Eskinazi-Sant'Anna & Tundisi, 1996 (tab.1,2); Hure & Krsinic, 1998 (p.103); Hopcroft & al., 1998 (tab.2); Lopes & al., 1998 (p.195); Suarez-Morales & Gasca, 1998 a (p.111); Lopes & al., 1999 (p.215, tab.1); Neumann-Leitao & al., 1999 (p.153, tab.2); Dalal & Goswami, 2001 (p.22, fig.2); Ara, 2001 b (p.121); Torres-Sorando & al., 2003 (p.157, population dynamics, growth, matrix model); Krumme & Liang, 2004 (p.407, tab.1); Ara, 2004 (p.179, figs. 3, 4, 5); Sterza & Fernades, 2006 (p.95, Table 1, occurrence); Morales-Ramirez & Suarez-Morales, 2008 (p.514); Miyashita & al., 2009 (p.815, Tabl.II); Mazzocchi & Di Capua, 2010 (p.428); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Costa R.G. da & al;, 2011 (p.364, Table 1, seasonal occurrence); Pinheiro & al., 2011 (p.425, abundance vs season); Magris & al., 2011 (p.260, abundance, interannual variability); Miyashita & al., 2012 (p.1557, Table 2: occurrence); Garbosa da Costa & al., 2013 (p.756, Table 1, abundance vs tide); Araujo & al., 2016 (p.1, Table 3, 5, abundance, %) ; Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Atique & al., 2017 (p.1, Table 1, fig.8). | | | | NZ: | 7 | | |
|
Carte de distribution de Oithona hebes par zones géographiques
|
| | | | | | | | | | Loc: | | | off Morocco, Brazil (S, Paranagua Bay, Vitoria Bay, Cananeia Lagoon, Mucuri estuary, Guarau estuary, do Pina estuary, NE, Bragança: Caeté Bay, off Natal, Ajuruteua Beach, Caeté Estuary, Amazon estuary), off Amazon, E Central America, Venezuela (brackish), Bahia de Mochima, Caribbean Colombia (Tayrona), Caibbean, Porto Rico, Jamaica, E Costa Rica, Texas (Laguna Madre), Ibero-moroccan Bay, Medit. (Alboran Sea, Banyuls, G. of Lion, Ligurian Sea, Tyrrhenian Sea, Malta, Adriatic Sea, Venice, W India (Mandovi & Zuari estuaries), Australia (Shark Bay) (in Kimmerer & al., 1985), Ecuador ( Guayaquil mouths), W Panama, Samoa Is. | | | | N: | 49 | | | | Lg.: | | | (46) F: 0,55; (179) F: 0,56-0,53; M: 0,54-0,48; (237) F: 0,55-0,60; M: 0,50-0,55; (449) F: 0,55; (635) F: 0,58-0,49; M: 0,5-0,47; (649) F: 0,55; (780) F: 0,61-0,47; M: 0,57-0,45; (880) F: 0,53-0,56; M: 0,48-0,54; {F: 0,47-0,61; M: 0,45-0,57}
(880): as O. ovalis) F: 0,58. | | | | Rem.: | épipélagique. Parfois dans les estuaires.
Les localisations en Méditerranée et en Australie nécessitent confirmation.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 12/02/2021 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 22 juin 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
