|
|
 |
Fiche d'espèce de Copépode |
|
|
Cyclopoida ( Ordre ) |
|
|
|
Oncaeidae ( Famille ) |
|
|
|
Oncaea ( Genre ) |
|
|
| |
Oncaea venusta Philippi, 1843 (F,M) | |
| | | | | | | Syn.: | Non Antaria gracilis Dana,1849; 1852;
Oncaea pyriformis Lubbock, 1860;
Antaria coerulescens Claus, 1866 (p.19);
Oncaea obtusa : Brady, 1883 (part., p.120, figs.F,M); Thompson, 1888 d (p.148); Kovalev & Shmeleva, 1982 (p.85);
Oncäa venusta : Giesbrecht, 1892 (p.590, 602, 774, figs.F, M); ? Razouls, 1972 (p.95, Annexe: p.111, figs.F,M); 1974 b (p.236, figs.F,M);
Oncaea praeclara Humes, 1988 (p.475, figs;F,M); Suarez-Morales & Gasca, 1998 a (p.112) | | | | Ref.: | | | Wheeler, 1901 (p.190, figs.F,M); Thompson & Scott, 1903 (p.239); A. Scott, 1909 (p.243, Rem.); Wolfenden, 1911 (p.362); Pesta, 1920 (p.652, fig.); Früchtl, 1924 b (p.88); Farran, 1929 (p.210, 284); Rose, 1929 (p.57); Wilson, 1932 a (p.353, figs.F,M); Rose, 1933 a (p.296, figs.F,M); Farran, 1929 (p.284, Rem.: 2 forms); Dakin & Colefax, 1933 (p.208); Farran, 1936 a (p.126); Mori, 1937 (1964) (p.119, figs.F,M); Dakin & Colefax, 1940 (p.116, figs.F,M); Lysholm & al., 1945 (p.43); Sewell, 1947 (p.263, Rem.: 2 formes); Sewell, 1951 (p.371, fig. juv.M, Rem.: parasites); Krishnaswamy, 1953 (p.68, Rem.); Marques, 1958 a (p.135); Chiba & al., 1957 (p.310); 1957 a (p.12); Tanaka, 1960 (p.71, Rem., large form.); Fish, 1962 (p.23); Fagetti, 1962 (p.45); Kasturirangan, 1963 (p.78, figs.F,M); Tanaka, 1964 (p.14); Vilela, 1965 (p.14); Saraswathy, 1966 (1967) (p.101); Owre & Foyo, 1967 (p.111, figs.F,M); Vilela, 1968 (p.31); Ramirez, 1969 (p.91, figs.F, Rem.); Corral Estrada, 1970 (p.216, Rem.); Shih & al., 1971 (p.56, 215); Björnberg, 1972 (p.89, figs.N, Rem.N, Juv.1-5); Marques, 1973 (p.247); Chen & al., 1974 (p.40, figs.F,M, p.75: Rem, forma major); Ferrari, 1975 (p.225, figs.F,M, Rem.); Marques, 1975 (p.51); 1976 (p.1000); Boxshall, 1977 a (p.124, figs.F,M, Rem. F: p.127: 3 formes); Dawson & Knatz, 1980 (p.9, 10, figs.F,M); Björnberg & al., 1981 (p.667, figs.F,M); Marques, 1982 (p.770); Malt, 1983 (p.454, fig.F, Rem.); 1983 a (p.7, figs.F,M, Rem.); J-s. Ho, 1984 a (p.41, figs.F, Rem.); Zheng Zhong & al., 1984 (1989) (p.262, figs.F,M); Huys Boxshall, 1991 (p.298, 300, 311, 314, figs.); Kim & al., 1993 (p.271); Lin & Nakamura, 1993 (p.448); Heron & Bradford-Grieve, 1995 (p.33, figs.F,M); Chihara & Murano, 1997 (p.981, Pl.223: F,M, Rem.:, Figs.371: left a,b = f.venella, right a, e = f.typica); Boxshall, 1998 (p.227); Böttger-Schnack, 2001 (p.30, Figs.F,M, 46, Rem., Biogéo.); Heron, 2002 (p.150, fig.F, tab.1, Rem.: p.153-154); Conway & al., 2003 (p.224, figs.F,M, Rem.); Boxshall & Halsey, 2004 (p.614: figs.); Böttger-Schnack & Huys, 2004 (p.1, Rem.: variant forms, size variants: small, medium, large); Avancini & al., 2006 (p.138, Pl. 106, figs.F,M, Rem.); Elvers & al., 2006 (p.503, phylogenetic relationships among different size groups); Wi & al., 2008 (p.184, figs.F,M, Rem.); Vives & Shmeleva, 2010 (p.311, figs.F,M, Rem.); Böttger-Schnack & Machida, 2011 (p.111, Table 1, fig.2, DNA sequences, phylogeny); Böttger-Schnack & Schnack, 2013 (p.4: Table 1, Rem.: s.str.-Group); Fuentes-Reinés & Suarez-Morales, 2017 (p.514, figs.F, Rem.) | 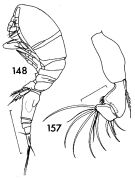 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.90, Lam. XVIII, figs.148, 157]. Female (from off Mar del Plata): 148, habitus (lateral left side); 157, A2. Scale bars in mm: 0.3 (148); 0.05 (157).
|
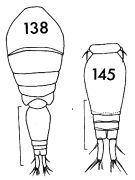 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.86, Lam. XVII, figs.138, 145]. Female (from off Mar del Plata): 138, habitus (dorsal); 145, urosome (dorsal). Scale bars in mm: 2 (138); 0.2 (145).
|
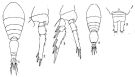 issued from : Q.-c Chen & S.-z. Zhang & C.-s. Zhu in Studia Marina Sinica, 1974, 9. [Pl.6, Figs.1-5]. Female (from off-shore Chekiang Province): 1, habitus (dorsal); 2, P3; 3, P4. Male: 4, habitus (dorsal); 5, posterior portion of abdomen.
|
 issued from : R. Böttger-Schnack in Bull. nat. Hist. Mus. Lond. (Zool.), 2001, 67 (1). [p.32, Fig.2]. Oncaea venusta f. typica. Female (from Red Sea): A, habitus (dorsal; lateral raised pore enlarged); B, idem (lateral left side); C, urosome (dorsal); D, idem (lateral left side); E, A1 (small sensory element arrowed); F, caudal ramus (dorsal; setae and numbered using Roman numerals); G, P6.
|
 issued from : R. Böttger-Schnack in Bull. nat. Hist. Mus. Lond. (Zool.), 2001, 67 (1). [p.33, Fig.3]. Oncaea venusta f. typica. Female: A, A2 (posterior; lateral elements are numbered using Roman numerals, distal elements indicated by capital letters); B, labrum (anterior, slit-like pores arrowed); C, idem (posterior); D, Md (showing individual elements, identified using capital letters); E, Mx1; F, Mx2; G, Mxp.
|
 issued from : R. Böttger-Schnack in Bull. nat. Hist. Mus. Lond. (Zool.), 2001, 67 (1). [p.34, Fig.4]. Oncaea venusta f. typica. Female: A, P1 (anterior) [a, 3rd endopod segment, showing aberrant spine number); B, P2 (anterior); C, P3 (anterior); D, P4 (anterior).
|
 issued from : R. Böttger-Schnack in Bull. nat. Hist. Mus. Lond. (Zool.), 2001, 67 (1). [p.36, Fig.6]. Oncaea venusta f. typica. Male (from Red Sea): A, habitus (dorsal; arrows indicating position of lateral raised pores); B, A1; C, Mxp (anterior); D, urosome (dorsal); E, idem (ventral); F, idem (lateral; spermatophores fylly developed); G, P5 (dorsal); H, A2 (posterior).
|
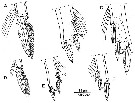 issued from : R. Böttger-Schnack in Bull. nat. Hist. Mus. Lond. (Zool.), 2001, 67 (1). [p.37, Fig.7]. Oncaea venusta f. typica. Male (from Red Sea): A, P1 (distal part of endopod); B, P2 (distal part of endopod); C, P3 (distal part of endopod). Oncaea venusta f. venella. Male (from Red Sea): D, P1 (distal part of endopod); E, P2 (distal part of endopod); F, P3 (distal part of endopod).
|
 issued from : R. Böttger-Schnack in Bull. nat. Hist. Mus. Lond. (Zool.), 2001, 67 (1). [p.51, Table 3]. Comparison of morphological characters of Oncaea venusta Gisbrecht from the Gulf of Naples with two forms, f. typica and f. venella from the Red Sea.
|
 issued from : R. Böttger-Schnack in Bull. nat. Hist. Mus. Lond. (Zool.), 2001, 67 (1). [p.50, Table 2]. Body length (mm) of Oncaea venusta f. typica and venella from the Red Sea..
|
 issued from : R. Böttger-Schnack & R. Huys in Hydrobiologia, 2004, 513. [p.3, Table 1]. Total body length (mm) of size variants of female in different areas of the Atlantic, Indian and Pacific Ocean. Size groups exhibiting a dorsal swelling on the P2-bearing somite are marked in bold. * = single specimen.
|
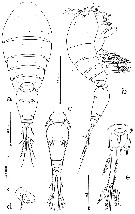 issued from : A.G. Humes in J. Plankton Res., 1988, 10 (3). [p.477, Fig.1]. As Oncaea praeclara. Female (from Galapagos Rift): a, habitus (dorsal); b, idem (lateral); c, urosome (dorsal); d, genital area (lateral); e, caudal ramus (dorsal). Nota: Ratio of length of prosome to that of urosome 1.49:1. Caudal ramus with fine setules along the inner margin, elongate, 125 microm. length, 23 microm. wide proximally (17 medially and 20 distally, ratio 6.7:1 based on medial width
|
 issued from : A.G. Humes in J. Plankton Res., 1988, 10 (3). [p.477, Fig.2]. As Oncaea praeclara. Female: a, urosome with egg sac (lateral); b, rostrum and labrum (ventral); c, A1 (postero-inner); d, A2 (antero-outer); e, Md (anterior view); f, paragnaths, Mx1 and labial area (ventral); g, Mx2 (antero-outer); h, Mxp (anterior); i, claw of Mxp (posterior).
|
 issued from : A.G. Humes in J. Plankton Res., 1988, 10 (3). [p.477, Fig.3]. As Oncaea praeclara. Female: a, area between Mxp and P1 (ventral); b, idem (lateral); c, P1 and intercoxal plate (anterior); d, P2 (anterior); e, P3 (anterior); e, P3; f, P4 (anterior); g, P5 (dorsal).
|
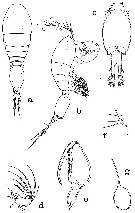 issued from : A.G. Humes in J. Plankton Res., 1988, 10 (3). [p.477, Fig.4]. As Oncaea praeclara. Male: a, habitus (dorsal); b, idem (lateral); c, urosome (ventral); d, 3rd segment of A2 (postero-inner); e, Mxp (anterior); g, spermatophore, partly extruded from male (ventral). Nota: P5 reduced to low ridge bearing 2 setae (lengths 23 and 36 microm.), and 1 adjacent (length 31 microm.P6 represented by posteroventral flap on genital segment bearing spiniform process but no setae
|
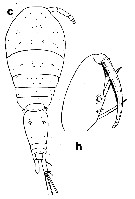 issued from : G.A. Boxshall in Brit. Mus. nat. Hist., Zool., 1977, 31 (3). [p.125, Fig.11, c, h]. As Oncaea venusta forma typica. Female (fro 18°N, 25°W): c, habitus (dorsal); h, Mxp.
|
 issued from : G.A. Boxshall in Brit. Mus. nat. Hist., Zool., 1977, 31 (3). [p.124]. Female: Armature formula of swimming legs P1 to P4. Roman numeral : spine; arabic numeral : seta.
|
 issued from : G.A. Boxshall in Brit. Mus. nat. Hist., Zool., 1977, 31 (3). [p.126, Fig.12, a-d]. Male: a, urosome (ventral); b, A2 (anterior); c, Mx1 (posterior); d, Mxp (anterior). Nota: Caudal rami about 1.8 times longer than anal somite and about 2.2 times longer than broad. A1 4-segmented. Mx1 bilobed; outer lobe with 4 setae, the outermost seta is slender and unarmed; inner lobe with 3 setae.
|
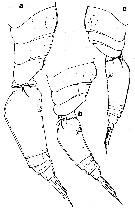 issued from : G.A. Heron in Hydrobiologia, 2002, 480. [p.153, Fig.6]. Female: pediger 2-4 and urosome (lateral). a, Oncaea venusta Philippi; b, Oncaea frosti Heron; c, Oncaea venella Farran. Nota: O. venusta is the largest of the three species. In preserved samples, it is rare to find either sex of the species where the urosome is not flexed at a 45° angle to the prosome, and the two postgenital segments and the anal seglment telescoped. Exoskeleton strongly chitinized and ornamentation conspicuous; pediger 2 without a conspicuous mid-dorsal dilationnin lateral view; caudal ramus length variable, slightly shorter or longer than the sum of the three preceding segments. Freshly collected specimens of O. venusta from the Gulf of naples and New Zeland areas both showed a purple-crimson shading on the widest portion of the prosome, posterior of genital segment, anterior of caudal rami, maxillipeds, and of the legs.
|
 issued from : G.A. Heron in Hydrobiologia, 2002, 480. [p.152, Table 1]. Length measurements (: body length, PL: prosome length, in mm) for adult females and males of Oncaea venusta, O. frosti and O. venella. Data are mean lengths (bold), number of specimens measured (parentheses), ranges, and standard error (se) at each locality; -, no specimens.
|
 issued from : J.H. Wi, H.-L Suh, H.S. Yang & H.Y. Soh in Ocean Sci. J., 2008, 43 (4). [p.185, Fig.1]. Female (from Yellow Sea, Korea waters): A-B, habitus (dorsal and lateral, respectively); C, A1; D, A2; E, Md ( with 6 elements indicated by small letters); F, Mx1; G, Mx2; H, Mxp; I, P5; J, P6. Nota: A1 6-segmented. Md: gnathobase with 5 elements (a-e); ventral element a shorter than ventral blade, with long, fine spinules along dorsal side; ventral blade b strong and extensive spiniform, with row of setules on posterior part; dorsal blade c strong and broad, with 3 dentiform processes along distal margin; 2 dorsal elements setiform, of which ventral one d shorter, flat and densely setose, dorsalmost one e longer and multipinnate. Mx1 single segmented, weakly bilobed: inner lobe (= praecoxal arthrite) with 3 elements and outer lobe with 4 elements (outermost one long spiniform having row of spinules). Mx2 2-segmented: syncoxa unarmed; allobasis produced distally into slightly curved claw bearing 2 rows of very strong spinules along medial margin; outer margin with strong seta extending almost to tip of allobasal claw, ornamented with few minute spinules distally; inner margin with slender pinnate seta and strong spine. Mxp 4-segmented, composed of syncoxa, basis extensive and robust, with 2 spiniform spinulose elements nearly equal in length and patches of long setules on inner margin; 1st endopodal segment without ornamentation; 2nd one with spinulose claw along concave margin, naked seta on outer proximal margin and unipectinate spine joined to inner margin. Proportional lenths (%) of urosomal segments and caudal rami 9.52 : 47.6 : 6.7 : 9.52 : 20.0. P5 with small plumose growing from lateral surface of somite, and small free exopod without ornamentation, exopod slightly longer than wide, bearing 2 naked setae. P6 expressed as operculum around each genital aperture with spine. Caudal rami about 3 times as long as wide with 6 elements.
|
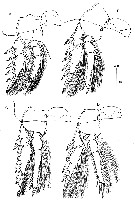 issued from : J.H. Wi, H.-L Suh, H.S. Yang & H.Y. Soh in Ocean Sci. J., 2008, 43 (4). [p.186, Fig.2]. Female: A-D, P1 to P4.
|
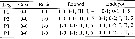 issued from : J.H. Wi, H.-L Suh, H.S. Yang & H.Y. Soh in Ocean Sci. J., 2008, 43 (4). [p.185, Fig.1]. Female: Armature formula, spines (Roman numerals) and setae (Arabic numerals).
|
 issued from : J.H. Wi, H.-L Suh, H.S. Yang & H.Y. Soh in Ocean Sci. J., 2008, 43 (4). [p.188, Fig.3]. Male: A-B, habitus (dorsal and lateral, respectively); C, A1; D, A2; E, Mxp; F-I, endopods of P1 to P4, respectively; J, P5. Nota: A1 4-segmented; distal segment corresponding to fused segments 4-6 of female; A2 coxobasis having naked and short seta at innerdistal corner; distal endopodal segment with seta III more stout than in female, seta IV spiniform and curved, both elements shorter than in female. P6 expressed as posterolateral flap closing off genital aperture on either side; covered by minute denticles.
|
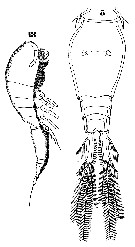 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 : 1-831, Atlas von 54 Tafeln. [Taf.47, Figs.5, 13]. As Oncäa venusta. Female: 5, habitus (dorsal); 13, same (lateral).
|
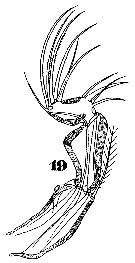 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.47, Fig.19]. As Oncäa venusta. Female: 19, A2 (posterior view).
|
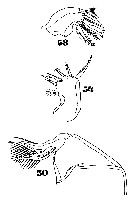 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19, Atlas von 54 Tafeln. [Taf.47, Figs.50, 54, 58]. As Oncäa venusta. Female: 50, Mx2; 54, Mx1; 58, Md.
|
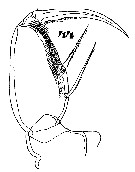 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19, Atlas von 54 Tafeln. [Taf.47, Fig.44]. As Oncäa venusta. Female: 44, Mxp.
|
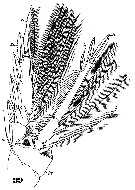 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.47, Fig.39]. As Oncäa venusta. Female: 39, P4 (posterior view).
|
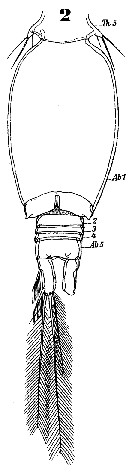 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.47, Fig.2]. As Oncäa venusta. Male: 2, urosome (ventral).
|
 issued from : C. Razouls in Th. Doc. Etat Fac. Sc. Paris VI, 1972, Annexe. [Fig.66]. Female (from Banyuls, G. of Lion): A, urosome; B, Mxp; C, A1; D, A2; E, P4; F, endopodite of P3; G, P1; H, P2.
|
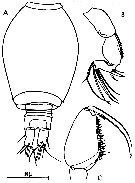 issued from : C. Razouls in Th. Doc. Etat Fac. Sc. Paris VI, 1972, Annexe. [Fig.67]. Male: A, urosome; B, A2; C, Mxp.
|
 Issued from : G.A. Heron & J.M. Bradford-Grieve in New Zealand Oceanogr. Inst Memoir 104. NIWA, 1995. [p.34, Fig.14, e-l] Female: e, habitus (lateral) [q]; f, same (dorsal) [q]; g, anterior of prosome (ventral) [r]; h, right A2 [s]; i, labrum (ventral) [u]; j, right Md [u]; k, left Mx1 [u]; l, right Mx2 [w]. Scale bars: See p.13, fig.2. letter in brackets.
|
 Issued from : G.A. Heron & J.M. Bradford-Grieve in New Zealand Oceanogr. Inst Memoir 104. NIWA, 1995. [p.35, Fig.15, a-j] Female: a, right Mxp [s]; b, P1 [y]; c, P2 [y]; d, P3 [y]; e, P4 [y]; f, P5 [s]. Male: g, habitus (lateral) [q]; h, same (dorsal) [q]; i, 3rd segment of right A2 [s]; j, left Mxp [s]. Scale bars: See p.13, fig.2. letter in brackets.
|
 Issued from : J.M. Fuentes-Reinés & E. Suarez-Morales in Check List, 2017, 13 (5). [p.515, Figs. 4, 5]. Oncaea venusta Female (from Rodadero Bay, N Colombia): 4, urosome (dorsal); 5, Anal somite and caudal rami (dorsal). Nota: Genital double-somite about 1.8 times as long as wide. - Anal somite with paired dorsal pore on posterior margin. - Caudal ramus about 3.5 times as long as wide.
| | | | | Ref. compl.: | | | Cleve, 1904 a (p.194); Pearson, 1906 (p.34, Rem.); Carl, 1907 (p.17); Rose, 1925 (p.152); Wilson, 1932 (p.50); 1942 a (p.199); Massuti Alzamora, 1942 (p.102, Rem.); Oliveira, 1945 (p.191); Wilson, 1950 (p.273); Sewell, 1948 (p.346, 461, 487); Yamazi, 1958 (p.154, Rem.); Deevey, 1960 (p.5, Table II, annual abundance) ; Ganapati & Shanthakumari, 1962 (p.10, 16); Duran, 1963 (p.25); Giron-Reguer, 1963 (p.58); V.N. Greze, 1963 a (tabl.2); Shmeleva, 1963 (p.141); Grice, 1963 a (p.496); Gaudy, 1963 (p.31, Rem.); Björnberg, 1963 (p.79, Rem.); De Decker, 1964 (p.16, 23, 30, 32); De Decker & Mombeck, 1964 (p.13); Bodo & al., 1965 (p.219, annual cycle); Anraku & Azeta, 1965 (p.13, Table 2, fish predator); Shmeleva, 1965 b (p.1350, lengths-volume -weight relation); Furuhashi, 1966 a (p.295, vertical distribution in Kuroshio region, Table 9); Neto & Paiva, 1966 (p.30, Table III, annual cycle); Mazza, 1966 (p.73); 1967 (p.365); Ehrhardt, 1967 (p.743, geographic distribution, Rem.); Séguin, 1968 (p.488); Evans, 1968 (p.14); Delalo, 1968 (p.139); Dowidar & El-Maghraby, 1970 (p.269); Deevey, 1971 (p.224); Gamulin, 1971 (p.382, tab.3); Binet & al., 1972 (p.71); Della Croce & al., 1972 (p.1, Rem.); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night, Table II: %, Table IV); Apostolopoulou, 1972 (p.329, 378); Björnberg, 1973 (p.361, 388); Guglielmo, 1973 (p.399); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Palet, 1975 (p.660); Vives & al., 1975 (p.56, tab.II); Peterson, 1975 (p.66 & suiv., abundance); Zalkina, 1977 (p.339, tab.1); Boxshall, 1977 b (p.546); Deevey & Brooks, 1977 (p.156, tab.2, Station "S"); Frontier, 1977 a (p.17); Binet, 1979 (p.400); Dessier, 1979 (p.132, 201, 207); Chen Q-c, 1980 (p.795); Vaissière & Séguin, 1980 (p.23, tab.2); Svetlichnyi, 1980 (p.28, Table 1, passive submersion); Star & Mullin, 1981 (p.1322, abundance); Gallo, 1981 (p.847); Vives, 1982 (p.295); Rudyakov, 1982 (p.208, Table 2); Arashkevich & al., 1982 (p.477, Table 2 cont., diet); Dessier, 1983 (p.89, Tableau 1, 2, Rem., %); Turner & Dagg, 1983 (p.17, 22); Gaudy & Boucker, 1983 (p.37, Table 1 3, 5, fig.1, 2, metabolism); Tremblay & Anderson, 1984 (p.7); Guangshan & Honglin, 1984 (p.118, tab.); Scotto di Carlo & al., 1984 (p.1044); Ho, 1984 a (p.23); Binet, 1984 (tab.3); 1985 (p.85, tab.3); Longhurst, 1985 (tab.2: as venustra); Regner, 1985 (p.11, Rem.: p.40); Sazhina, 1985 a (p.491, tab.3); Jansa, 1985 (p.108, Tabl.I, II, III, IV); Petipa & Borichenko, 1985 (tab.2); Turner, 1986 (p.289, fig. 2- 6: pellet-content); Valentin & al., 1986 (p.117, Table V); Sarkar & al., 1986 (p.178); M. Lefèvre, 1986 (p.33); Renon, 1987 (tab.2); Dessier, 1988 (tabl.1); Jimenez-Perez & Lara-Lara, 1988; Williams D. & al., 1988 (p.580); Lozano Soldevilla & al., 1988 (p.61); Hernandez-Trujillo, 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Herman, 1989 (p.247); Othman & al., 1990 (p.561, 564, Table 1); Hirakawa & al., 1990 (tab.3); Yoo, 1991 (tab.1); Hirakawa, 1991 (p.376: fig.2); Hernandez-Trujillo, 1991 (1993) (tab.I); Seguin & al., 1993 (p.23); Böttger-Schnack, 1994 (p.277); Hwang & Turner, 1995 (p.207, Table 1, 2, fig. 1, 2, 3: swimming behaviour); Palomares Garcia & Vera, 1995 (tab.1); Webber & Roff, 1995 (tab.1); Hirakawa & al., 1995 (tab.2); Shih & Young, 1995 (p.77); Böttger-Schnack, 1995 (p.92); 1996 (p.1086); Webber & al., 1996 (tab.1); Kotani & al., 1996 (tab.2); Suarez-Morales & Gasca, 1997 (p.1525); Park & Choi, 1997 (Appendix); Go & al., 1998 (p.475, Table 1, feeding behavior); Hure & Krsinic, 1998 (p.87, 104); Krsinic, 1998 (p.1051); Noda & al., 1998 (p.55, Table 3, occurrence); Lopes & al., 1998 (p.195, tab.2); Hopcroft & al., 1998 (tab.2); Alvarez-Cadena & al., 1998 (t.1,2,3,4); Hsieh & Chiu, 1998 (tab.2); Suarez-Morales & Gasca, 1998 a (p.112); Neumann-Leitao & al., 1999 (p.153, tab.2); Siokou-Frangou, 1999 (p.479); Dolganova & al., 1999 (p.13, tab.1); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); Lopes & al., 1999 (p.215, tab.1); Voronina & Kolosova, 1999 (p.72); Harvey & al., 1999 (p.1, 49: Appendix 5, in ballast water vessel); Lapernat, 2000 (tabl.3, 4); Razouls & al., 2000 (p.343, tab. 5, Appendix); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); Ueda & al., 2000 (tab.1); Fernandez-Alamo & al., 2000 (p.1139, Appendix); Suarez-Morales & al., 2000 (p.751, tab.1); Seridji & Hafferssas, 2000 (tab.1); Lopez-Salgado & al., 2000 (tab.1); Alvarez-Silva & Gomez-Aguirre, 2000 (p.163: tab.2); Moraitou- Apostolopoulou & al., 2000 (tab.I); Böttger-Schnack & al., 2001 (p.1029, tab. 1, 2); Dalal & Goswami, 2001 (p.22, fig.2); Lo & al., 2001 (1139, tab.I); Krsinic & Grbec, 2002 (p.127, tab.1); Zerouali & Melhaoui, 2002 (p.91, Tableau I); Sameoto & al., 2002 (p.13); Vukanic, 2003 (p.139, tab.1); Hsieh & al., 2004 (p.398, tab.1, p.399, tab.2); Rezai al., 2004 (p.490, tab.2, Rem., p.495, tab.8); Lo & al.*, 2004 (p.218, tab.1, fig.6); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Lo & al., 2004 (p.468, tab.2); Chang & Fang, 2004 (p.456, tab.1); Daly Yahia & al., 2004 (p.366, fig.4); Lan & al., 2004 (p.332, tab.1, tab.2); Kang & al., 2004 (p.1524, fig.9); Satapoomin & al., 2004 (p.109, tab.6); Nishibe & Ikeda, 2004 (p.931, Tab. 2, 5); Lo & al., 2004 (p.89, tab.1); Vukanic & Vukanic, 2004 (p.9, tab. 3); Kazmi, 2004 (p.231, Rem.: p.232); Dias Bonecker, 2005 (p.100 + poster); Alvarez-Silva & al., 2005 (p.39); Berasategui & al., 2005 (p.485, tab.1); Prusova & Smith, 2005 (p.76, 78); Berasategui & al., 2006 (p.485: fig.2); Zuo & al., 2006 (p.164: tab.1); Rakhesh & al., 2006 (p.93, Table 2, spatial distribution); Hwang & al., 2006 (p.943, tabl. I); Sterza & Fernades, 2006 (p.95, Table 1, occurrence); Dias & Araujo, 2006 (p.89, Rem., chart); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño); Durbin & Casas, 2006 (p.2537, Table 2a, 2b); Fang & al., 2006 (p.224, metals concentration); Khelifi-Touhami & al., 2007 (p.327, Table 1); Hwang & al., 2007 (p.25); Tseng L.-C. & al., 2008 (p.153, fig.5, Table 2, occurrence vs geographic distribution, indicator species); Tseng L.-C. & al., 2008 (p.46, table 2: abundance vs moonsons, table 3: indicator species, fig.8); Jitlang & al., 2008 (p.65, Table 1); Rakhesh & al., 2008 (p.154, Table 5: abundance vs stations); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); McKinnon & al., 2008 (p.844: Tab.1, fig.7: 2 forms typica & "medium form"); Humphrey, 2008 (p.85: Appendix A); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.525); Lopez Ibarra, 2008 (p.1, Table 1, fig.11: abundance); Fernandes, 2008 (p.465, Tabl.2); Pagano, 2009 (p.117); Ohtsuka & al., 2008 (p.115, Table 5); Chiba & al., 2009 (p.1846, Table 1, occurrence vs temperature change); C.-Y. Lee & al., 2009 (p.151, Tab.2); Miyashita & al., 2009 (p.815, Tabl.II); Böttger-Schnack & Schnack, 2009 (p.131, Table 3, 4, Rem.); Tseng & al., 2009 (p.327, fig.5, feeding); Licandro & Icardi, 2009 (p.17, Table 4); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Nishibe & al., 2009 (p.491, Table 1: seasonal abundance, 3 forms: %); C.E. Morales & al., 2010 (p.158, Table 1); Hafferssas & Seridji, 2010 (p.353, Table 2); Lidvanov & al., 2010 (p.356, Table 3); Zhang G.-T. & al., 2010 (p.492, Table 1); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Hidalgo & al., 2010 (p.2089, fig.2, 4, Table 2, cluster analysis); Dias & al., 2010 (p.230, Table 1); Mazzocchi & Di Capua, 2010 (p.429); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Hafferssas & al., 2010 (p.1281, Table III, abundance vs spatial distribution); Fazeli & al., 2010 (p.153, Table 1); W.-B. Chang & al., 2010 (p.735, Table 2, 3, 4, fig.5, abundance); Hsiao & al., 2010 (p.179, Table I, II, III, trace metal concentration); Xu & Gao, 2011 (p.514, figs.3, 4, Table 2: optimal salinity); Hsiao S.H. & al., 2011 (p.475, 481: indicator species, Appendix I); Hsiao & al., 2011 (p.232, Table 1: abundance, %, Table 2: trace metal concentration, vs transect); Hsiao & al., 2011 (p.317, Table 2, 3, fig.6, indicator of seasonal change); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Chen H; & al., 2011 (p.84, spatial & temporal variations); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Tutasi & al., 2011 (p.791, Table 1, 3, fig.7, abundance distribution vs La Niña event, Rem. p.796, fig.11); Shiganova & al., 2012 (p.61, Table 4); Uysal & Shmeleva, 2012 (p.909, Table I); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Zizah & al., 2012 (p.79, Tableau I, Rem.: p.86); Salah S. & al., 2012 (p.155, Tableau 1); Jose & al., 2012 (p.20, fig.3 a, b, c:, % vs monsoon); Miyashita & al., 2012 (p.1557, Table 2: occurrence); Johan & al., 2012 (2013) (p.1, Table 1); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Tseng & al., 2012 (p.621, Table 3: abundance, indicator species); Hidalgo & al., 2012 (p.134, Table 2, 3, figs.6, 8, occurrence vs hydrology) ; Gubanova & al., 2013 (in press, p.4, Table 2); Palomares-Garcia & al., 2013 (p.1009, Table I, abundance vs environmental factors); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Hsiao & Fang, 2013 (p.175, Hg bioaccumulation); Melo Jùnior & al., 2013 (p.363, Table 2, egg production); Tseng & al., 2013 (p.507, seasonal abundance); Tseng & al., 2013 a (p.1, Table 3, abundance); Rakhesh & al., 2013 (p.7, Table 1, 4, abundance vs stations); Varadharajan & Soundarapandian, 2013 (p.2: occurrence vs stations); Jagadeesan & al., 2013 (p.27, Table 3, 5, fig.11, seasonal variation); Anjusha & al., 2013 (p.40, Table 3, abundance & feeding behavior); Alvarez-Silva & Torres-Alvarado, 2013 (p.241, Table1: seasonal abundance); Sano & al., 2013 (p.11, Table 2, feeding habits); Lidvanov & al., 2013 (p.290, Table 2, % composition); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Lopez-Ibarra & al., 2014 (p.453, fig.6, biogeographical affinity); Zakaria, 2014 (p.3, Table 1, abundance vs. 1960-2000); Zaafa & al., 2014 (p.67, Table I, occurrence); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production, Table 4: % vs. season); Rojas-Herrera & al., 2016 (p.40, Table 2: temporal abundance); Araujo & al., 2016 (p.1, Table 3, abundance, %); Rosa J.C.L. & al., 2019 (p.71, Rem.); Zakaria & al., 2016 (p.1, Table 1); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Ben Ltaief & al., 2017 (p.1, Table III, Summer relative abundance); El Arraj & al., 2017 (p.272, table 1, distribution vs. seasonal variability);Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence) ; Ohtsuka & Nishida, 2017 (p.565, Table 22.1); Bonecker C.T. & al., 2017 (p.247, fig.8: abundance day/night vs. vertical types of mass water); El Arraj & al., 2017 (p.272, table 1, 2, spatial distribution); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence); Dias & al., 2018 (p.1, Tables 2, 4, 5: vertical distribution, abundance vs. season); Palomares-Garcia & al., 2018 (p.178, Table 1: occurrence); Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions). | | | | NZ: | 23 | | |
|
Carte de distribution de Oncaea venusta par zones géographiques
|
| | | | | | | | | | | | | | | | | | 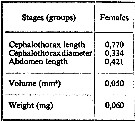 issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 42]. Oncaea venusta (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 42]. Oncaea venusta (from South Adriatic).
Dimensions, volume and Weight wet. Means for 50-60 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
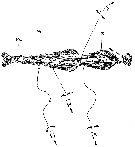 issued from : Y.-B. Go, B.-C. Oh & M. Terazaki in J. Mar. Syst., 1998, 15. [p.480, Fig.4]. issued from : Y.-B. Go, B.-C. Oh & M. Terazaki in J. Mar. Syst., 1998, 15. [p.480, Fig.4].
Attacking behaviors of Oncaea spp. on the body of Sagitta (Chaetognatha). FVL: head trunk in front fins; VG: ventral ganglion; CS: caudal septum
Nota: Direct underwater observations with SCUBA were performed at night. The attack behavior of Oncaea started mostly below the stationary Sagitta in the field. The attack distance at which Oncaea spp. approached Sagitta from below was about 5-6 cm; the attacking behavior from the upper side of Sagitta was observed only occasionally, and the attack distance was about 1-3 cm. This behaviour suggests that Oncaea is not a touch feeder. The attachment sites of Oncaea on the body of Sagitta spp. were for a total of 320 individuals: ventral side 56.3 %, lateral side 32.8 % and dorsal side 10.9 %.
Oncaea crept directly to the chaetognath tail or the head region using their second antennae, and pierced the body of chaetognaths with the long claw of the maxilliped, and moved maxillae and mandibles.
The chaetognaths did not move at all when this copepod was crawling on the body. |
 Issued from : P. Tutasi, S. Palma & M. Caceres in Scienc. Mar., 2011, 75 (4). [p.798, Fig. 7a] Issued from : P. Tutasi, S. Palma & M. Caceres in Scienc. Mar., 2011, 75 (4). [p.798, Fig. 7a]
Geographic distribution of Oncaea venusta in September and October 2001, associated with the weak La Niña event of 2001. |
 Issued from : S.-H. Hsiao, J.-S. Hwang & T.-H. Fang in Crustaceana, 2010, 83 (2). [p.184, Table I]. Issued from : S.-H. Hsiao, J.-S. Hwang & T.-H. Fang in Crustaceana, 2010, 83 (2). [p.184, Table I].
Copepods collected from the sea around northern Taiwan and from southern East China Sea extending to the Okinawa Trough, to study possible spatial heterogeneity. The content of the same metal shows considerable variation both intra- and inter-specifically. The metal concentrations in males are higher than in females. Copepod metal quota display spatial variation: coastal water > southern East China Sea > Kuroshio water, suggesting that the metal contents of copepods are influenced by the water quality of their marine environment. |
 Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6]. Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6].
Variations in the most abundant copepod species (mean ± SE) along the transect in the boundary waters between the northern part Taiwan Strait and the East China Sea i March (black bar) and October (grey bar) 2005 (Mann-Whitney U test, sig. *P <0.05.
See drawings of Hydrological conditions and superficial marine currents in Calanus sinicus. |
 Issued from : A.C.T. Bonecker, A. V. de Araujo, C.O. Dias, M.M.S. Castro, P.F. Carvalho, R.M. Lopes & S.C.L. Bonecker in A.P.C. Falcao & D.L. Moreira (ed.) Ambiente pelagico caracterizaçao ambiental regional de Bacia de Campos, Atlantico Sudoeste. Rio de Janeiro. Elsevier. 2017, v.5, p.247-281. [p.260, Fig.8] Issued from : A.C.T. Bonecker, A. V. de Araujo, C.O. Dias, M.M.S. Castro, P.F. Carvalho, R.M. Lopes & S.C.L. Bonecker in A.P.C. Falcao & D.L. Moreira (ed.) Ambiente pelagico caracterizaçao ambiental regional de Bacia de Campos, Atlantico Sudoeste. Rio de Janeiro. Elsevier. 2017, v.5, p.247-281. [p.260, Fig.8]
Abundance (ind. m-3) sampled during night and day into four water masses in the Campos Basin (Brazil).
AT: Tropical water; ACAS: Central water of South Atlantic; AIA: Intermediate water of Antarctica; ACS: Deep water Circumpolar water superior.
Sampling at station c (22°54'30''S, 40°43'W), depth 1900 m (end of the continental slope).
Nota: On 117 taxa, with 81 species identified, this species is one of the most abundant (219,09 ind.m-3), following Farranula gracilisClausocalanus furcatus. |
 Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.209: Table 1, p.210: Table 2]. Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.209: Table 1, p.210: Table 2].
Table 1: Experimental protocol.
Table 2:Swimming versus non-swimming behaviour, and behavioural transitions per minute. Means (bold type), ranges, and standard deviations (SD). Number of repeated observations = 500.
Copepods collected in surface net from waters near Pitou (NE coast Taiwan: 25°N, 122°E) in daylight in July 1993.
Compare with Oithona nana, O. similis, Macrosetella gracilis, Temora turbinatam> and Undinula vulgaris var. taiwanicus by the same authors. |
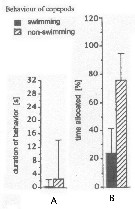 Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.211: Figs.1, 2]. Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.211: Figs.1, 2].
Fig.1: A, Mean durations periods (seconds) of swimming versus non swimming behaviour by Oncaea venusta from north-east coast of Taiwan (near Pitou) in daylight in July 1993.
Fig.2: B, mean percentages of time allocated to swimming versus non-swimming behaviour by Oncaea venusta. |
| | | | Loc: | | | Cosmopolite (equatorial, tropical, sub-tropical, temperate). From North Pacific, also : Bering Sea, Gulf of Alaska; from South : SE Australia, New Zealand. From the north Atlantic it seems with difficulty reach the latitude 45°, although noted from East Nova Scotia (in Sameoto & al., 2002), from Labrador and north Hudson Bay (in Wilson, 1936 d); From the south of South Africa and Buenos Aires, Brazil (S, off Rio de Janeiro, Campos Basin, Vitoria Bay, Ubatuba, off Macaé, Paranagua estuary, Camamu, off Natal), in Antarctic (Weddell Sea in Voronina & Kolosova (1999), sub-Antarctic (Indiian, SE Pacif.), Arctic. (continent), also: Hawaii (Kaneohe Bay), Clipperton Is., Galapagos Rift, E Pacif. Rise, Baja California (Bahia Magdalena, La Paz), G. of California (Guaymas Basin), Coyuca lagoon (Guerrero), Acapulco Bay, Caribbean Colombia, Rodadero Bay, W Costa Rica, G. of Mexico, off Mississipi River mouth, off Bermuda (Station "S"), Delaware Bay (outside), Bahia Cupica (Colombi), Peru, Chile (N, Concepcion), Australia (North West Cape), E Korea, Cheju Island, China Seas (Yellow Sea, East China Sea, South China Sea, Changjiang River estuary), off SW Taiwan, Taiwan Strait, Taiwan (S, NE, Pitou, E: Kuroshio & Oyashio Currents), Kueishan Is., Korea Strait, Japan ( Kuchinoerabu Is., Sagami Bay, Tosa Bay, Tokyo Bay, Seto Inland Sea), Malaysia (Sarawak: Bintulu coast), Ambon Bay, , SE India (Mallipayyinam-Manamelkudi, Mediterranean Sea (Black Sea included), G. of Guinea, off Lagos, Red Sea | | | | N: | 514 ? (probably less because inadequacy précision with O. venella, if it is maintened) | | | | Lg.: | | | (34) F: 1,33-0,84; (35) F: 1,16-1,08; M: 0,81-0,78; (45) F: 1,27-1,1; M: 0,95-0,8; (46) [Médit.] F: 1,27-1,1; M: 0,95-0,8; [Atlant.] F: 1,33-0,85; (52) F: 1,206-0,996; (66) F: 1,39-1,13; M: 1,07-0,74; (91) F: 1,28-1; M: 1-0,8; (104) F: 1,25; M: 1; (109) F: 1,35-1,2; M: 1,1-0,9; (114) F: 1,25-1,15; 0,96-0,94; (139) F: 1,13-0,98; M: 1,08-0,96; (180) F: 1,25-1,05; M: 0,73; 0,67; (254) F: 1,7-1,25; (327) F: 1,29-0,99; M: 0,92-0,79; (334) F: 1,27-1,1; M: 1-0,7; (336) F: 1,5-0,97; M: 0,85-0,61; (373) M: 0,86; (449) F: 1,27-1,1; M: 1-0,7; (530) F: 1,2; M: 0,9; (531) F: 1; M: 0,8; (618) F: 1,37-1,3; (651) F: 1,39-0,92; M: 0,96-0,88; (687) [G1] F: 0,99-0,92; M: 0,63-0,57; [G2] F: 1,2-1,1; M: 0,86-0,76; (699) F: 1,01-1,30; M: 0,91-0,98; (785) F: 1,33-0,85; (786) F: 1,5-0,99; M: 0,87; (864) F: 1,1-1,3; (865) F: 1,27; (920) F: 1,16; (991) F: 1-1,28; M: 0,7-1; (1020) F: 0,96-1,2; M: 0,78- 0,88; (1125) F: 0,9-1,0; (1313) F: 0,94-0,965; {F: 0,84-1,70; M: 0,57-1,10}
Nota: Fuentes-Reinés & Suarez-Morales (2017, p.516) underline that the species O. venusta is known for its high variability in total body length (ranging from 0.75 to 1.4 mm in the female and from 0.55 to 1.98 mm in the male/ Based on its size range the species has been categorized into 3 groups. Molecular studies could differentiate 4 generic clades where the small and large size groups were separated genetically and both could be deemed as distinct species (see Elvers & al., 2006). Therefore, some authors prefer to designate the group with large size as O. venusta §0.960-1.260 mm) and the smaller forms as O. venella (0.800-0.830 mm) | | | | Rem.: | Epi à mésopélagique, aussi démersal and semi-parasitic.
Sampling depth (Antarct., sub-Antarct.) : 400 m.
Cette espèce présente 2 formes (sous-espèces ?) qui sont caractérisées par leurs dimensions. Sewell (1947) suggère aussi que leur saison de ponte est différente.
Heron (2002) considère les deux variétés O. venusta typica et O. venusta venella comme des espèces (voir remarques à Oncaea venella et frosti), position contestée par Böttger-Schnack & Huys (2004).
Humes (1988) décrit O. praeclara prélevé à très grande profondeur (2000 à 2635 m) sur des sites d'hydrothermalisme du Pacifique est.
L'abondance donnée par Dias & al. (2018) doit être très sous-estimée du fait de lusage d'un filet de maille > 200 µm.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 28/10/2022 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 22 mai 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
