 Liste des auteurs étudiés
Liste des auteurs étudiés
Deevey, 1960 (p.5, Table II, fig.8: annual abundance); H. Schulz, 1961 (p.57); Engel, 1962 (p.252, american occurrences); Northcote & al., 1964 (p.1069, Table II); Dimov, 1964 a (p.31); Heinle, 1969 (p.186, temperature effect); 1969 a (p.150, Table 1, culture); Katona, 1970 (p.373, generation times, sex ratios, reproduction, survival vs. temperature); 1973 (p.574, Table 1, 2, fig.1, sex pheromones); Eriksson, 1973 a (p.95, fig.8, area abundance distibution); Katona, 1975 (p.89, Table 1I, II, figs., copulation, egg production); Bradley, 1975 (p.26, t° & Sal. effects); Heinle & Flemer, 1975 (p.235, carbon requirements); Richman & al., 1977, (p.69, grazing: particle selection); Allan & al., 1977 (p.317, grazing); Heinle & al., 1977 (p.341, feeding, egg production); Harris & al., 1977 (p.187, polluant effect); Berdugo & al., 1977 (p.138, reproduction vs pollution); Ott & al., 1978 (p.49, toxic effect); Bradley, 1978 (p.177, temperature acclimatation); Knatz, 1978 (p.68, Table 1, fig.3, abundance, %); Collins & Williams, 1981 (p.273, monthly distribution-salinity); Grice & Marcus, 1981 (p.125, Dormant eggs, Rem.: p.135); Burkill & Kendall, 1982 (p.21, production); Boak & Goulder, 1983 (p.139, Rem.: bacterioplankton); Jacoby & Youngbluth, 1983 (p.84, Table 4, Rem: mating); Barthel, 1983 (p.269, filtration, ingestion, respiratory rates); Tremblay & Anderson, 1984 (p.6,
Rem.); Roddie & al., 1984 (p.191, S-T tolerance & osmoregulation); Tremblay & Anderson, 1984 (p.6), Rem.); Williams & Collins, 1985 (p.28); Bradley, 1986 (p.561, genetic/temperature/salinity); Kankaala & Johansson, 1986 (p.1027, length-biomass); Heckman, 1986 (p.176, anadromus migration); Vuorinen & Ranta, 1987 (tab.2, 4); Sellner & Bundy, 1987 (p.1435, turbidity effects); Giguère & al., 1989 (p.522, Table 1, formaldehyde preservation); Castel & Feurtet, 1989 (p.577, population dynamic); Stoecker & McDowell Capuzzo, 1990 (p.891, feeding); Jouffre & al., 1991 (p.489, lagoon); Naess, 1991 (p.261, fig.6, 7, resting egg); Bradley & Davis, 1991 (p.321, survival vs. temperature & genetic); Bradley & al., 1992 (p.3, molecular biology); Hough & Naylor, 1992 (p.27, endogenous rhythms); Viitasalo, 1992 (tab.2); Hirche, 1992 (p.395, Table 1, egg production, k-sreategy); Ban, 1992 (p.361, diapause, egg production vs environmental factors); Ban & Minoda, 1992 (p.51, hatching of diapause eggs); Pietsch, 1993 (p.215, mortality rates); Escaravage & Soetaert, 1993 (p.201, development vs. t°, secondary production); Castel, 1993 (p.145, long-term annual distribution); Ban, 1994 (p.721, temperature & food concentration effects); Ban & Minoda, 1994 (p.185, diapause egg production); Marcus & al., 1994 (p.154, Rem. p.157); Sellner & al., 1994 (p.249); Hoffmeyer, 1994 (p.303); Pietsch, 1995 (p.127, production); Irigoien & al., 1996 (p.163, gut clearance rate); Madhupratap & al., 1996 (p.77, Table 2: resting eggs); Ban S, Burns C. & al., 1997 (p.287, fig.1, Table 1, 2, feeding, reproduction); Cordell & al., 1997 (p.7, % change vs. 1980-1997); Suarez-Morales & Gasca, 1998 a (p.111); Mauchline, 1998 (tab.8, 18, 21, 26, 33, 35, 40, 45, 46, 47, 48, 51, 56, 58, 61, 62, 63, 65); Hairston & Bohonak, 1998 (p.23, Table 3); Viitasalo & al., 1998 (p.129, behaviour vs. predator): Gasparini & Castel, 1999 (p.877, diet); Gasparini & al., 1999 (p.195, egg production); Harvey & al., 1999 (p.1, 49: Appendix 5, in ballast water vessel); Lee, 1999 (p.1423, invasive); Ueda & al., 2000 (tab.1); Peitsch & al., 2000 (p.175, seasonal-long term abundances vs. transect); Burdloff & al., 2000 (p.165, egg production); Holmes, 2001 (p.27); Miramand & al., 2001 (p.1056, matals concentration); Kimmel & Bradley, 2001 (p.135, protein expression vs Salinity-temperature variation); Mouny & Dauvin, 2002 (p.13, fig.3, 5, 6, Table 4); Tackx & al., 2003 (p.305, feeding); Titelman & Kiørboe, 2003 a (p.137, nauplius behaviour); Lee & Petersen, 2003 (p.296, acclimatization); Forget-Leray & al., 2005 (p.268, toxicity vs development); Cordell & al., 2007 (p.218, fig.7); David & al., 2007 (p.377); 2007a (p.429, figs.2, 4, 5, Table 2, 3, 4, 5, 6); Cailleaud & al., 2007 (p.270, contamination); 2007 a (p.281); Devreker & al.,2007 (p.1117, post-embryonic development vs t°, Sal.); 2008 (p.1329), tidal effects); Höök & al., 2008 (p.29); Darnis & al., 2008 (p.994, Table 1); Suarez-Morales & al., 2008 (p.679, occurrence); Devreker & al., 2008 (p.1329, abundance vs tidal estuary); Devreker & al., 2009 (p.113, reproduction vs t° & Sal.); Beyrend-Dur & al., 2009 (p.713, table II, development time, reproduction, vs salinity); Devreker, 2009 (p.113, reproduction vs temperature & salinity); Brylinski, 2009 (p.253: Tab.1, p.255: Rem.); Cailleaud & al., 2009 (p.64, contaminant effects); Telesh & al., 2009 (p.16, fig.2.1, p.18: Table 2.1); Dur & al., 2009 (p.1073, reproduction); Seuront, 2010 (p.1223: salinity effect); Souissi & al., 2010 (p.1227: salinity effect); Michalec & al., 2010 (p.805); Mialet & al., 2010 (p.116); Devreker & al., 2010 (p.245); Modéran & al., 2010 (p.219, seasonal distribution); Dvoretsky & Dvoretsky, 2010 (p.991, Table 2); Bollens & al., 2011 (p.1358, Table III, fig.8); Strasser & al., 2011 (p.1210, fecundity model vs environment); Schmitt & al., 2011 (p.773, Rem.: behavior); Steele C.W. & Scarfe, 2011 (p.142, model courtship/copulation vs population density); Cailleaud & al., 2011 (p.228, behavior); Dur & al., 2011 (p.197, behavior); Elliot & Tang, 2011 (p.1039, spatial & temporal distribution); Mahjoub & al., 2011 (p.1095, vulnerability to predator); Sakaguchi & al., 2011 (p.18, Table 2, occurrences); Sopanen & al., 2011 (p.1564, mortality vs algal toxicity); Cailleaud & al., 2011 (p.226, contaminant bioconcentration); Glippa & al., 2011 (p.255, resting eggs); Postel, 2012 (p.327, Table 1); Delpy & al., 2012 (p.1921, Table 2); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Stefanova & al., 2012 (p.403, Table 2, interannual abundance); Drillet & al., 2012 (p.155, Table 1, culture); Bollens & al., 2012 (p.101, Table 1, species competition); Vehmaa & al., 2012 (p.351, reproduction vs algal composition); Madéran & al., 2012 (p.126, feeding adaptation vs turbidity); Devreker & al., 2012 (p.72, development, egg production); Holliland & al., 2012 (p.298, diel vertical migration); Arula & al., 2012 (p.33, Table 2: interannual abundance); Loick-Wilde & al., 2012 (p.199, feeding, N2 incorporation); Gusmao & al., 2013 (p.279, Table 3, 4, sex-specific predation by fish, fig.3: sex ratio vs temperature); Dur & al., 2013 (p.21, population dynamics model); Gissi & al., 2013 (p.86, Table 3, copper toxicity); Gorokhova & al., 2013 (p.1, pigmentation vs predation); Souissi & al., 2013 (p.38, behavior vs ciliate epibionts); Seuront, 2013 (p.724, mate-feeding behaviour); Michalec & al., 2013 (p.129, behavior vs aquatic toxics); Lesueur & al., 2013 (p.60, survival, growth vs toxicity); Lloyd & al., 2013 (p.299, egg production); Bradley C.J. & al., 2013 (p.49, swimming behavior); Setälä & al., 2014 (p.77, table 1, fig.1, microplastic ingestion); Kimmerer & al., 2014 (p.722, reproduction , growth vs food); Buskey & al., 2017 (p.754, amyelinate fiber vs escape response); Hansen B.W. & al., 2017 (p.984, pH effects); Baumgartner & Tarrant, 2017 (p.387, resting eggs, Rem.); Richirt & al., 2019 (p.3, Table 1, fig.2, 3, 4, 5, abundance changes vs years 1998-2014, table 2: diversity index)


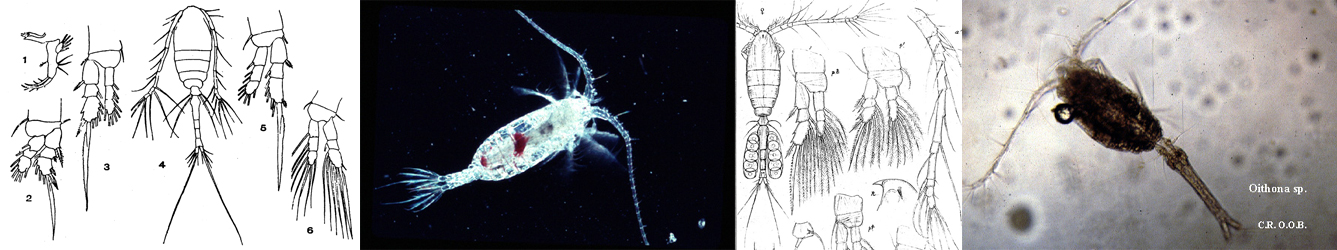

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}