 Liste des auteurs étudiés
Liste des auteurs étudiés
Wolfenden, 1904 (p.112, Descr.F, figs.F); A. Scott, 1909 (p.13, figs.F, Rem.); Wolfenden, 1911 (p.196, figs.F,M); With, 1915 (p.41, figs.F, Rem.); Lysholm & Nordgaard, 1921 (p.8); Farran, 1926 (p.230); Jespersen, 1934 (p.45); Farran, 1939 (p.360); Jespersen, 1940 (p.9); Vervoort, 1946 (p.49, figs. juv.,
Rem.); Sewell, 1947 (p.25,
Rem.: abnormality, fig.F); 1948 (p.555, Rem.); [Non Tanaka, 1956 (p.262, figs.F)]; Vervoort, 1957 (p.32,
Rem.); 1963 b (p.86, Rem.); Paiva, 1963 (p.16, figs.F); Grice, 1963 a (p.495); Mazza, 1966 (p.69); Tanaka & Omori, 1967 (p.241); Owre & Foyo, 1967 (p.34, figs.F,M); Vinogradov, 1968 (1970) (p.268); Guérédrat, 1969 (p.65,
Rem.); Lee & al., 1971 (p.1151); Silas, 1972 (p.645); [Non Brodsky & al., 1983 (p.191, figs.F, M, Rem.F)]; Roe, 1984 (p.356); Tremblay & Anderson, 1984 (p.4, Rem.); Guangshan & Honglin, 1984 (p.118, tab.); Fleminger, 1985 (p.276, 284, Table 1, fig.M)); Nishida, 1989 (p.173, table 2, 3, dorsal hump); Michel, 1994 (p.177,
Rem.); [Non Bradfod-Grieve, 1994 (fig.18, Gigs. F,M)]; Chihara & Murano, 1997 (p.834, Pl.126: F,M); Bradford-Grieve & al., 1999 (p.877, 905, figs.F,M); Boxshall & Halsey, 2004 (p.141: figs.); Vives & Shmeleva, 2007 (p.912, figs.F,M, Rem.); Andronov, 2014 (p.33, fig.M); Bradford-Grieve & al., 2017 (p.30, Descr. F,M, NVI, copepodite II, figs. F,M, N6, juv.II ,Rem., phylogeny)


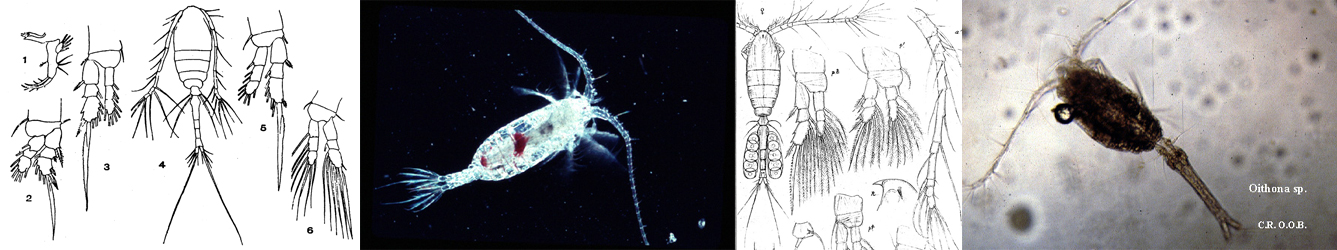

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}