|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Arietelloidea ( Superfamille ) |
|
|
|
Augaptilidae ( Famille ) |
|
|
|
Haloptilus ( Genre ) |
|
|
| |
Haloptilus longicornis (Claus, 1863) (F,M) | |
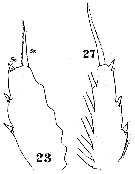
| | | | | | | Syn.: | Hemicalanus longicornis Claus, 1863 (p.179, fig.F); Brady, 1883 (p.44); Thompson, 1888 d (p.139); Giesbrecht, 1892 (p.384, 398, 772, figs.F,M); T. Scott, 1894 b (p.32);
? Haloptilus setuliger Tanaka, 1964 b (p.40, Descr.F, figs.F);
Haloptilus longicirrus : Bradford, 1970 a (p.359, figs.F);
Haloptilus longicornis s.l. : Brinton & al., 1986 (p.228, Table 1) | | | | Ref.: | | | Giesbrecht & Schmeil, 1898 (p.118, Rem. F,M); Sars, 1902 (1903) (p.121, figs.F); Farran, 1908 b (p.67); A. Scott, 1909 (p.140, Rem.); Wolfenden, 1911 (p.323); Pesta, 1920 (p.534); Sars, 1925 (p.240); Farran, 1926 (p.286); 1929 (p.209, 269); Rose, 1933 a (p.210, figs.F,M); Farran, 1936 a (p.113); Mori, 1937 (1964) (p.75, figs.F); Lysholm & al., 1945 (p.37); Brodsky, 1950 (1967) (p.362, figs.F, Rem.M); Chiba & al., 1957 (p.308); 1957 a (p.11); Grice, 1962 (p.223, Rem.F); Tanaka, 1964 b (p.39, figs.F,M); Chen & Zhang, 1965 (p.86, figs.F,M); Vervoort, 1965 (p.123, 125, Rem.); Saraswathy, 1966 (1967) (p.80); Owre & Foyo, 1967 (p.81, figs.F,M, Rem.); Park, 1968 (p.561, Rem.F); Ramirez, 1969 (p.79, fig.F); Park, 1970 (p.535, figs.F, Rem.); Corral Estrada, 1970 (p.191, figs.F, Rem.); Tanaka & Omori, 1971 (p.253); Matthews, 1972 (p.50, Rem.); Björnberg, 1972 (p.50, fig.N, Rem.N); Kos, 1972 (Vol.I, figs.F,M, Rem.); Razouls, 1972 (p.95, Annexe: p.79); Marques, 1976 (p.994); Björnberg & al., 1981 (p.653, figs.F,M); Zheng & al., 1982 (p.58, figs.F); Ali-Khan & Ali-Khan, 1984 (p.308, figs.F); Park, 1988 (p.18, Descr.F, figs.F, Rem.); Zheng Zhong & al., 1984 (1989) (p.250, figs.F); Chihara & Murano, 1997 (p.728, Pl.60,62: F,M); Bradford-Grieve & al., 1999 (p.883, 941, figs.F,M); Bradford-Grieve,1999 b (p.61, figs.F,M, Rem., figs.172, 190); Conway & al., 2003 (p.84, figs.F,M, Rem.); Mulyadi, 2004 (p.185, figs.F, Rem.); Vives & Shmeleva, 2007 (p.242, figs.F,M, Rem.); Blanco-Bercial & al., 2011 (p.103, Table 1, Biol. mol, phylogeny) | 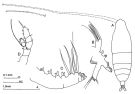 issued from : J.M. Bradford-Grieve in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). NIWA Biodiversity Memoir, 111, 1999. [p.61, Fig.32]. Female: A, habitus (dorsal); B, Mx1; C, Mx2; D, P5.
|
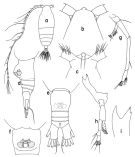 issued from : T. Park in Antarct. Res. Ser. Washington, 1988, 47. [p.19, Fig.11]. Female: a, habitus (dorsal); b, forehead (dorsal); c, idem (ventral); d, urosome (lateral left side); e, urosome (dorsal); f, genital segment (ventral); g, A2; h, Md; i, Md (cutting edge). Nota: The forehead has an anterior protrusion which in dorsal view is clearly set off. Ventrally, the spermathecae are very close to each other and to the genital opening. The caudal ramus is about 1.5 times as long as it is wide. Mx1 usually 4 setae on the basis.
|
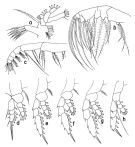 issued from : T. Park in Antarct. Res. Ser. Washington, 1988, 47. [p.20, Fig.12]. Female: a, Mx1; b, Mx2; c, Mxp; d, P1 (anterior); e, P2 (anterior); f, P3 (anterior); g, P4 (posterior); h, P5 (posterior).
|
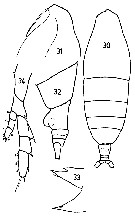
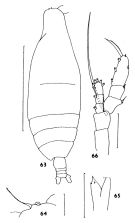 issued from : J.M. Bradford in N.Z. Jl Mar. Freshw. Res., 1970, 4 (4). [p.360, Figs 63-65]. As Haloptilus longicirris. Female (off Kaikoura, New Zealand): 63, habitus (dorsal); 64, forehead (dorsal); 65, Md (cutting edge); 66, P5. Scale bars represent 1.0 mm in 63 and 0.1 mm in 64-66. Nota: A1 is twice the length of the body. The two distal teeth of the mandibular blade are not equal. The spine on the 2nd basipod of P5 is over twice the length of the endopod.
|
 issued from : G.O. Sars in An Account of the Crustacea of Norway. Vol. IV. Copepoda Calanoida. Published by the Bergen Museum, 1903. [Pl.LXXXII]. Female. Nota: R = rostrum (ventral view); M = Md; m = Mx1.
|
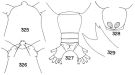 Issued from : T. S. Park in Bull. Mar. Sc., 1970, 20 (2). [p.534, Figs.325-329]. Female (from Caribbean Sea & G. of Mexico): 325, forehead (dorsal); 326, idem (ventral); 327, urosome (dorsal); 328, genital segment (ventral); 329, Md (biting edge of the mandibular blade). Npta: Forehead with a protrusion which in dorsal view is clearly set off. Genital segment wider than long, with two spermathecae located close to each other. Caudal rami about 1.5 times as long as wide. Basis of Mx1 usually with 4 setae.
|
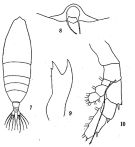 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.34, è-10]. Female (E China Sea): 7, habitus (dorsal); 8, rostrum (ventral view); 9, Md; 10, right P5 (posterior).
|
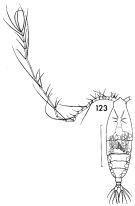 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.78, Lam. XV, figs.123]. Female (from off Mar del Plata): 123, habitus (dorsal). Scale bars in mm: 1
|
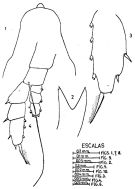 issued from : J. Corral Estrada in Tesis Doct., Univ. Madrid, A-129, Sec. Biologicas, 1970. [Lam.50, figs.1-4]. Female (from Canarias Is.): 1, forehead (dorsal); 2, Md (cutting edge); 3, exopodal segment 3 of P4; 4, P5.
|
 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.59, Fig.33]. Female: a, habitus (dorsal); b, forehead with rostrum (ventral view); c, urosome (lateral); d, A2; e, Md; f, Md (biting edge); g, P1; h, P5. Scale bar in mm.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.38, Figs.5-14]. Female: 5, P5; 6, Mxp; 7, Mx2; 8, habitus (dorsal); 9, Md; 10, P1; 11, P2; 12, P4; 13, A2; 14, P3.
|
 issued from : S. Ali-Khan & J. Ali-Khan in Crustaceana, 1984, 47 (3). [p;308, Figs.30-34]. Female (from 24°09'N, 64°27'E): 30, habitus (dorsal); 31, forehead (lateral); 32, last thoracic segment and urosome (lateral); 33, Md (biting edge); 34, P5.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.27 , Figs.3, 29 ]. Hemicalanus longicornis. Male: 3, grasping A1 (distal portion); 29, P5 (posterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.27 , Figs.8, 9, 10 ]. Hemicalanus longicornis. Female: 8, Mx2 (posterior view); 9, A2 (anterior view); 10, Mx1 (posterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.27 , Fig.33]. Hemicalanus longicornis. Female: 33, Md (mandibular blade).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.27 , Figs.23, 27]. Hemicalanus longicornis. Female: 23, exopodite 3 of P4; 27, exopodite 3 of P2.
|
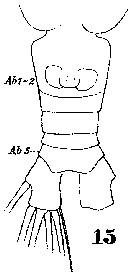 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.42 , Fig.15]. As Hemicalanus longicornis. Female: 15, urosome (ventral). Ab 1-2 = genital double-somite; Ab 5 = anal somite
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.42 , Fig.29]. As Hemicalanus longicornis. Male: 29, A2.
|
 Issued from : G.S. Brady in Rep. Scient. Results Voy. Challenger, Zool., 1883, 8 (23). [Pl. IX, Figs.1-7]. Male: 1, habitus (dorsal); 2, A2; 3, Md; 4, Mx1; 5, Mx2; 6, Mxp. Female: 7, P5.
|
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.80, Fig.530]. Female: 530, Md (gnathobase).
|
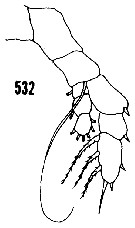 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.80, Fig.532]. Female: 532, P5.
|
 issued from : P.E. Lapernat & C. Razouls in Vie Milieu, 2002, 52 (1). [p.23, Pl. III, fig.5]. Biting edge of Md gnathobase female (from off Malta, Mediterranean Sea). The Itoh's index value = 4091 (number of teeth: 2).
|
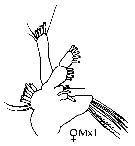 issued from : J.M. Bradford-Grieve, E.L. Markhaseva & C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, Edit. D. Boltovskoy. 1999. Vol.2. Copepoda. [p.1043, Fig. 7.272]. Female: Mx1. Noya: Exopodite of Mx1 with 7 setae.
|
 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.105, Fig.38]. Female (from 07°29'S, 121°05'E): a, habitus (dorsal); b-c, forehead (dorsal and ventral, respectively); d, urosome (dorsal); e, posterior thoracic segment and urosome (lateral left side); f, P5. Nota: Forehead with a characteristic knob-like projection. A1 about twice longer than body. P5 almost symmetrical, distal seta on inner exopod segment 3 not longer than other setae.
| | | | | Ref. compl.: | | | Pearson, 1906 (p.27); Rose, 1924 d (p.480); Wilson, 1942 a (p.188); Massuti Alzamora, 1942 (p.111); Sewell, 1948 (p.350, 504, 509, 514, 516, 526, 547, 549, 558, 567); C.B. Wilson, 1950 (p.236); Krishnaswamy, 1953 (p.126); Yamazi, 1958 (p.150, Rem.); Heinrich, 1960 (p.36); Gaudy, 1962 (p.93, 99, Rem.: p.112) ; Fagetti, 1962 (p.31); Ganapati & Shanthakumari, 1962 (p.8, 15); V.N. Greze, 1963 a (tabl.2); Shmeleva, 1963 (p.141); Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); Duran, 1963 (p.23); Gaudy, 1963 (p.27, Rem.); Grice, 1963 a (p.496); Björnberg, 1963 (p.54, Rem.); Deevey, 1964 (p.589, Table 3, lengths variation); Unterüberbacher, 1964 (p.30); De Decker & Mombeck, 1964 (p.12); Shmeleva, 1965 b (p.1350, lengths-volume-weight relation); Grice & Hulsemann, 1965 (p.224); Pavlova, 1966 (p.44); Furuhashi, 1966 a (p.295, vertical distribution in Kuroshio region, Table 9); Mazza, 1966 (p.72); 1967 (p.367); Ehrhardt, 1967 (p.741, geographic distribution, Rem.); Grice & Hulsemann, 1967 (p.19); Fleminger, 1967 a (tabl.1); Vinogradov, 1968 (1970) (p.79, 82, 91, 277); Delalo, 1968 (p.138); De Decker, 1968 (p.45); Evans, 1968 (p.13); Berdugo & Kimor, 1968 (p.447); Dowidar & El-Maghraby, 1970 (p.268); Park, 1970 (p.478); Deevey, 1971 (p.224); Gamulin, 1971 (p.381, tab.2); Binet & al., 1972 (p.69); Roe, 1972 (p.277, tabl.1, tabl.2); 1972 c (p.1033); Apostolopoulou, 1972 (p.328, 362); Heinrich, 1973 (p.95); Björnberg, 1973 (p.347, 386); Harding, 1974 (p.141, tab. 3, gut contents); de Bovée, 1974 (p.109, 124); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Vives & al., 1975 (p.50, tab.II, III); Deevey & Brooks, 1977 (p.156, tab.2, Station "S"); Tranter, 1977 (p.596); Carter, 1977 (1978) (p.36); Dessier, 1979 (p.206); Vaissière & Séguin, 1980 (p.23, tab.2); Svetlichnyi, 1980 (p.28, passive submersion); Madhupratap & al., 1981 (p.266, fig.1q: abundance vs. geographic transect); Vives, 1982 (p.293); Rudyakov, 1982 (p.208, Table 2); Kovalev & Shmeleva, 1982 (p.84); Scotto di Carlo & Ianora, 1983 (p.150); Guangshan & Honglin, 1984 (p.118, tab.); Roe, 1984 (p.358); De Decker, 1984 (p.317); Cummings, 1984 (p.163, Table 2); Stephen, 1984 (p.161, Distribution vs thermocline & geographic); Scotto di Carlo & al., 1984 (1041); Longhurst, 1985 (tab.2); Almeida Prado Por, 1985 (p.250); Regner, 1985 (p.11, Rem.: p.36); Brenning, 1985 a (p.28, Table 2); 1986 (p.13, Rem.); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Madhupratap & Haridas, 1986 (p.105, tab.1); Ambler & Miller, 1987 (tab.2, 3, 4, 5); Lozano Soldevilla & al., 1988 (p.60); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Heinrich, 1990 (p.18); Suarez & al., 1990 (tab.2); Madhupratap & Haridas, 1990 (p.305, fig.3: vertical distribution night/day; fig.7: cluster); Scotto di Carlo & al., 1991 (p.270); Hattori, 1991 (tab.1, Appendix); Yoo, 1991 (tab.1); Suarez & Gasca, 1991 (tab.2); Hernandez-Trujillo, 1991 (1993) (tab.I); Huntley & Lopez, 1992 (p.201, Table B1, growth rate, temperature-dependent production); Suarez, 1992 (App.1); Seguin & al., 1993 (p.23); Heinrich, 1995 (tab.1); Shih & Young, 1995 (p.68); Webber & Roff, 1995 a (p.481, table 1, 2, 3, biomass, production); Padmavati & Goswami, 1996 a (p.85, fig.3, vertical distribution); Hure & Krsinic, 1998 (p.69, 102, 112); Gilabert & Moreno, 1998 (tab.1, 2); Suarez-Morales, 1998 (p.345, Table 1); Suarez-Morales & Gasca, 1998 a (p108); Mauchline, 1998 (tab.58, 64); Siokou-Frangou, 1999 (p.478); Lopes & al., 1999 (p.215, tab.1); Lapernat, 1999 (p.9, 55); Razouls & al., 2000 (p.343, tab. 5, Appendix); Fernandez-Alamo & al., 2000 (p.1139, Appendix); Seridji & Hafferssas, 2000 (tab.1); Lopez-Salgado & al., 2000 (tab.1); Lapernat & Razouls, 2001 (p.123, tab.1); Madhupratap & al., 2001 (p. 1345, vertical distribution vs. O2, figs.4, 5: clusters, p.1353); Holmes, 2001 (p.13); Beaugrand & al., 2002 (p.179, figs.5, 6); Vukanic, 2003 (p.139, tab.1); Lo & al., 2004 (p.89, tab.1); Vukanic & Vukanic, 2004 (p.9, tab. 2, 3); Shimode & al., 2005 (p.113 + poster); Berasategui & al., 2005 (p.313, fig.2); Kazmi, 2004 (p.227); Zuo & al., 2006 (p.163: tab.1); Isari & al., 2006 (p.241, tab.II); Dias & Araujo, 2006 (p.30, Rem., chart); Lavaniegos & Jiménez-Pérez, 2006 (p.145, tab.2, 4, Rem.); Zervoudaki & al., 2006 (p.149, Table I); Koppelmann & Weikert, 2007 (p.266: tab.3); Hwang & al., 2007 (p.23); Dur & al., 2007 (p.197, Table IV); Jitlang & al., 2008 (p.65, Table 1); McKinnon & al., 2008 (p.843: Tab.I, p.848: Tab. IV); Morales-Ramirez & Suarez-Morales, 2008 (p.513, 517); Fernandes, 2008 (p.465, Tabl.2); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Raybaud & al., 2008 (p.1765, Table A1); Lan Y.C. & al., 2008 (p.61, Table 1, % vs stations); Tseng L.-C. & al., 2008 (p.153, Table 2, fig.5, occurrence vs geographic distribution); C.-Y. Lee & al., 2009 (p.151, Tab.2); Galbraith, 2009 (pers. comm.); Licandro & Icardi, 2009 (p.17, Table 4); Brugnano & al., 2010 (p.312, Table 3, fig.8); Hafferssas & Seridji, 2010 (p.353, Table 3); Williamson & McGowan, 2010 (p.273, Table 3, Pacific central gyres: N and S); Cornils & al., 2010 (p.2076, Table 3); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Dias & al., 2010 (p.230, Table 1); Mazzocchi & Di Capua, 2010 (p.424); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); W.-B. Chang & al., 2010 (p.735, Table 2, 4, abundance); Dvoretsky & Dvoretsky, 2010 (p.991, Table 2); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Tutasi & al., 2011 (p.791, Table 2, abundance distribution vs La Niña event); Uysal & Shmeleva, 2012 (p.909, Table I); Miloslavic & al., 2012 (p.165, Table 2, transect distribution); Dorgham & al., 2012 (p.473, Table 4: abundance vs season); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Belmonte & al., 2013 (p.222, Table 2, abundance vs stations); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Peijnenburg & Goetze, 2013 (p.2765, genetic data); Norton, 2013 (p.1fig.1: geographic distribution, figs.2, 3, 4: genetic structures); Lidvanov & al., 2013 (p.290, Table 2, % composition); Siokou & al., 2013 (p.1313, fig.4, 8, biomass, vertical distribution); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Mazzocchi & al., 2014 (p.64, Table 3, 4, 5, spatial & seasonal composition %); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production); Zakaria & al., 2016 (p.1, Table 1); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); El Arraj & al., 2017 (p.272, table 2); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Belmonte, 2018 (p.273, Table I: Italian zones); Hure M. & al., 2018 (p.1, Table 1: abundance, % composition, Rem.: p.12); Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions). | | | | NZ: | 23 | | |
|
Carte de distribution de Haloptilus longicornis par zones géographiques
|
| | | | | | | | | | | | | | | 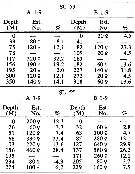 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.81, Table 37]. issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.81, Table 37].
Vertical distribution of H. longicornis at the ''40-Mile station in the Florida Current (± 25°35'N, 79°27'W.
SL 53 : 18 V 1958; SL 55: 21 VII 1958; A = day; B = night.
Nota: Roehr & Moore (1965) diagrammed its average noon and midnight distribution in the upper 350 m in the Florida Straits and gave its 30 p.cent level at 175 m at midday and 184 m at night. They interpreted this as showing a downward movement at night, i.e, a reverse migration. |
 issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 33 ]. Haloptilus longicornis (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 33 ]. Haloptilus longicornis (from South Adriatic).
Dimensions, volume and Weight wet. Means for 50-60 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
 issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.30]. issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.30].
Chart of occurrence in Brazilian waters (sampling between 22°-23° S).
Nota: sampling only 1 specimen. |
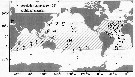 Issued from : E.L. Norton in Master of Sc. Oceanogr., Hawaii Univ., 2013. [p.36, fig.1]. Issued from : E.L. Norton in Master of Sc. Oceanogr., Hawaii Univ., 2013. [p.36, fig.1].
Collection sites for population included in the sudy of gene flow in the species Haloptilus longicornis.
Samples with n > 25 individuals (closed symbols) are included in all analyses. Samples with n < 25 individuals (open symbols) in the tropical Atlantic were included only in statistical parsimony analyses (fig.2). Collection sites are numbered as in Table 1. Hashed region indicates the apporoximate global biogeographic distribution of H. longicornis. A question mark indicated ocean region in which the species has been reported to occur, but considered uncertain.
In this study was investigated genetic connectivity among populations of this mesopelagic copepod, using the mitochondrial marker, cytochrome oxidase subunit II (mtCOII, 43 locations, N = 1059), highly significant genetic structure was detected among ocean basins, with the exception of a lack between South Atlantic and Indian Ocean populations.
Genetic breaks were also detected among subtropical gyre populations within the Atlantic and Pacific Oceans in this species.
The results indicate that the equatorial region may be poor quality habitat for this species and serve as a biophysical dispersal barrier to migrants traveling among subtropical gyre habitats.
In order to better understand contemporary dispersal patterns and the mechanism underlying this dispersal barrier in the Atlantic Ocean, the author simulated particle dispersal for 30 years using the Simple Ocean Data Assimilation (SODA) model coupled with a particle-tracking scheme.
The results suggest that the dispersal barrier' across the equatorial region must be biophysical in nature, and that it likely prevents contemporary dispersal among subtropical gyre populations. Therefore, populations of H. longicornis in the subtropical gyres of the northern and southern Atlantic are demographically as well as genetically isolated, and may evolve independently in response to local selective pressures. |
 Issued from : G. Deevey in J. mar. biol. Ass. U.K., 1964, 44. [p.596, Fig.3, A] Issued from : G. Deevey in J. mar. biol. Ass. U.K., 1964, 44. [p.596, Fig.3, A]
Histogram showing the length distribution, plotted as numbers measured of females of Haloptilus longicornis from 1958-1959, at Station "S' (32°8' N, 64°42' W), 15 miles south-east of Brermuda.
Samples by plankton net, oblique hauls from 500 m to the surface |
 Issued from : G. Deevey in J. mar. biol. Ass. U.K., 1964, 44. [p.597, Table 3] Issued from : G. Deevey in J. mar. biol. Ass. U.K., 1964, 44. [p.597, Table 3]
Mean cephalothorax lengths and numbers measured of females.
See in Pleuromamma abdominalis for yhe mean surface temperature and Chlorophyll a of the month preceding each measurement at Station 'S' (SE of Bermuda).
Correlation coefficients for mean length with the temperature and chlorophyll of the preceding month were calculated. No correlation was found between the lengths and either the temperature or the chlorophyll. The variations in length noted are not related to the total phytoplankton cycle or to seasonal temperature changes. |
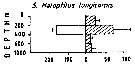 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.310, Fig.3]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.310, Fig.3].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 5- Haloptilus longicornis.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1].
Length-weight regression of Haloptilus longicornis in oceanic waters of Discovery Bay (Jamaica) in 60- and 200 m vertical hauls, using 200- and 64-µm mesh aperture of plankton nets, from June 1989 to July 1991 (monthly samples). |
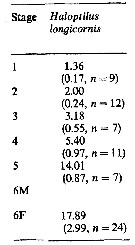 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 2]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 2].
Mean biomass (µg) of copepodite stages for Haloptilus longicornis from Discovery Bay (Jamaica. (SD: Standard deviation and n number of individuals measured. |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 3]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 3].
Stage weight (In mmax mmin-1) for Haloptilus longicornis from Discovery Bay (Jamaica), calculated from mean biomass of each stage. |
| | | | Loc: | | | Cosmopolite (tropical, temperate): Antarct. (Indian), South Africa (E & W), off Tristan da Cunha, Namibia, off Ascension Is. (SW & S), off S St. Helena Is., G. of Guinea, Ivorian shelf, S Cape Verde Is., Morocco-Mauritania, Canary Is., off Madeira, off Portugal, off W Cabo Finisterre, Bay of Biscay, Argentina, off Trinidade Is., Brazil (off Rio de Janeiro, Campos Basin, off Cabo de Sao Tomé, off Macaé, Amazon), Caribbean Colombia, Caribbean Sea, Jamaica, E Costa Rica, Yucatan, G. of Mexico, Florida, Sargasso Sea, off Bermuda (Station "S"), off E Cape Cod, off W Ireland, Porcupine Bank, off S Iceland, Barents Sea (Rare ?), Ibero-moroccan Bay, Medit. (Alboran Sea, NW Basin, Banyuls, Marseille (rare), Monaco, Ligurian Sea, Tyrrhenian Sea, Messina, Gulf of Taranto, Taranto, Malta, Adriatic Sea, Ionian Sea, Aegean Sea, Lebanon Basin, W Egyptian coast, Alexandria), G. of Elat, off Sharm El-Sheikh, Red Sea, G. of Aden, Arabian Sea, Pakistan, Indian, W India (Goa - Gujarat), E India (Lawson's Bay, Madras), Bay of Bengal, off Madagascar (S, SW), Nosy Bé, Natal, W Australia, North West Cape, Indonesia (Flores Sea, SW Celebes), Philippines, China Seas (Yellow Sea, East China Sea, South China Sea), Taiwan (SW, S, N: Mienhua Canyon, NW), S Korea, Korea Strait, Tanabe Bay, off SE Japan, Tokyo Bay, off Sanriku, Kuroshio zone, Pacif. (W equatorial), Pacific (central gyres: N and S), Australia (Great Barrier), New Zealand (N North Island, off E North Island, NE), Tasman Sea, Sw Pacif. (sub-Antarctic), off NE Hawaii, California, W Baja California, Gulf of California, G. of Tehuantepec, off W Guatemala, W Costa Rica, Pacif. (SE tropical), Galapagos-Ecuador, Peru, Chile | | | | N: | 226 | | | | Lg.: | | | (16) F: 2,23-1,94; (22) F: 2,5-2,1; M: 1,2; (34) F: 2,45-1,98; (35) F: 2,16-2,04; (38) F: 2,07-1,98; (46) F: 2,5-2,1; M: 1,18; (65) F: 2,15; (69) F: 2,25-2,14; M: 1,16; (72) F: 2,44-2; (88) F: 2,16-1,94; M: 1,24; (90) F: 2,63-2,1; (101) F: 1,9-1,77; (199) F: 2,58-1,82; M: 1,37; (237) F: 2,1; (254): F: ±2,5; (290) F: 2; (313) F: 2; (315) F: 2,04; (340) F: 2,2-2,1; (449) F: 2,5-2,1; M: 1,18; (530) F: 2; (795) F: 1,4; (866) F: 1,72-2,63; (909) F: 1,95-2,16; (920) F: 2,23; (991) F: 1,95-2,5; M: 1,18-1,24; (991) F: 1,95-2,5; M: 1,18-1,24; (1023) F: 2,17; (1108) F: 2,16; {F: 1,40-2,63; M: 1,16-1,37} | | | | Rem.: | épi-bathypélagique.
Sampling depth (Antarct.): 2000 m.
Owre & Foyo (1967) signalent que cette espèce présente une migration nycthémérale inverse.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 24/10/2022 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2025. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 02 décembre 2025] © copyright 2005-2025 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
