|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Calanoidea ( Superfamille ) |
|
|
|
Calanidae ( Famille ) |
|
|
|
Calanus ( Genre ) |
|
|
| |
Calanus chilensis Brodsky, 1959 (F,M) | |
| | | | | | | Ref.: | | | Brodsky, 1959 a (p.1543, figs.F,M); 1961 (p.8, figs.F,M); Kun, 1969 (p.997, figs.F) Heinrich, 1971 (p.292, Rem.); Brodsky, 1972 (1975) (p.9, 68, 82, 119, figs.); Bradford & Jillett, 1974 (p.6, fig.F); Arcos, 1975 (p.7, figs.F); Vyshkvartzeva, 1977 a (p.97, figs.); Fleminger, 1985 (p.275, 285, Table 1, 4, Rem.: A1); Bradford, 1988 (p.74, 76, Rem.); van der Spoel & Heyman, 1983 (p.62, fig.79); Marin & al., 1994 (p.75, Rem.M); Hill & al., 2001 (p.279, fig.2: phylogénie); Ferrari & Dahms, 2007 (p.35, Rem. N) | 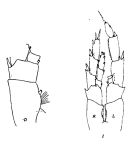 issued from : J.M. Bradford & J.B. Jillett in Crustaceana, 1974, 27 (1). [p.14, Fig.2,D,I]. Female: D, P2. Male: I, P5.
|
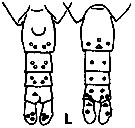 issued from K. Hulsemann in Invert. Taxon., 1994, 8. [p.1477, Fig.28, L]. Female: L, urosome (left: ventral); right: dorsal). Pore signature schematic by pooled samples (symbols are considerably larger than pores): Filled circle: 100 % presence; open circle: 95-99 % presence; triangle: 50-89 % presence. n = 21.
|
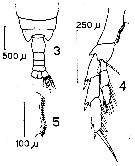 issued from : D.F.R. Arcos in Gayana, Zool., 1975, 32. [Lam.I, Figs.3-5]. Female (from Bahia de Concepcion, Chile): 3, posterior part of cephalothorax and urosome (dorsal); 4, P5; 5, inner margin of basipodite 1 of P5.
|
 issued from : K.A. Brodsky in Zool. Zh., 1959, 38, 10. [p.1541, Fig.3, 12]. Calanus chilensis Female (from 33°03'S, 72°10'W): 12, coxopodite inner edge of P5. Nota: Dentate plate on coxopodite of P5 has teeth of triangular shape with spaces between the teeth in distal part, spaces being narrower in proximal part of inner edge of this segment. Number of teeth 18-20. Curve present in middle part of teeth line but not pronounced. Distal segment of endopodite of P5 has 5 bristles. Relation of breadth of prosome to length 1 ; 2.8. A1 longer than body by 3-4 segments.
|
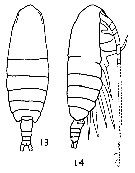 Issued from : K. Brodsky in Inform. Ser. 33. New Zealand Depart. Sc. Indust. Res., 1961. [p.9, Pl. I, fig. 13 and 14]. Female (from Valaparaiso, Chile): 13, habitus (dorsal); 14, same (lateral). Nota: Proportion wide to length of cephalothorax 1 : 2.8. Last thoracic segment with short, disvergent distal corners. A1 longer than the body of 3-4 segments.
|
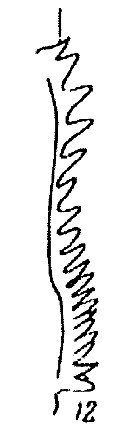
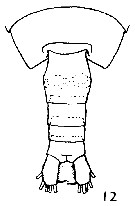 Issued from : K. Brodsky in Inform. Ser. 33. New Zealand Depart. Sc. Indust. Res., 1961. [p.10, Pl. 2, fig. 12]. Female (from Valparaiso, Chile): 12, last thoracic segment and urosome (dorsal). Nota; Abdomen relatively thicker and not as short as that of the other Calanus species.
|
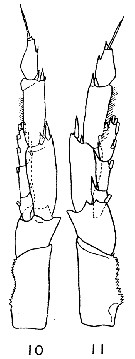 Issued from : K. Brodsky in Inform. Ser. 33. New Zealand Depart. Sc. Indust. Res., 1961. [p.13, Pl. 4, fig. 10 and 11]. Male (from Chile): Left leg of P5 (10; from Talcahuano; 11: from Valparaiso). Nota: P5 with relatively short, thick exopodites segments in the left leg. Endopodite of the left leg reaches as far as 1/3 or 1/2 the length of the 2nd exopodal segment of the same leg. Serrate plate of basipod with 17-18 teeth; distal teeth larger than proximal ones. 3rd exopodite segment of the left leg shorter and broader than that of Calanus australis. The 3rd endopodite segment of P5 has 6 setae.
| | | | | Ref. compl.: | | | Arcos, 1976 (p.85, Rem.: p.89, Table II); Judkins, 1980 (p.475, fig.5: vertical distribution vs O2 minimum); Herman & Mitchell, 1981 (p.739, Table 2, length-volume); Herman & Sameoto, 1981 (p.228, Table 1, 2, abundance); Herman A.W., 1984 (p.131, Rem.: p.133); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Peterson W.T. & Bellantoni, 1987 (p.411, Table I, figs.10, 11, egg production vs total chlorophyll concentration); Marin & al., 1994 (p.75, Rem.); Escribano & Rodriguez, 1994 (p.289); Ban S, Burns C. & al., 1997 (p.287, Table 1, 2, feeding, reproduction); Escribano & al., 1998 (p.19, development, growth vs. temperature); Gonzalez & Marin, 1998 (p.109); Huggett & Richardson, 2000 (p.1843, tab.2); Escribano & Hidalgo, 2000 (p.283, tab.2); 2000 a (p.1867, interannual abundance, growth); Hidalgo & Escribano, 2001 (p.157, fig.4); Giraldo & al., 2016 (p.1307, temperature effects); Poulet & al., 2007 (p.187, reproduction vs diatom concentration); Castro & al., 2007 (p.486), Table 1, 2, 3, 4, fig.3, 5); Morales C.E. & al., 2007 (p.452, fig.8, Rem.: p.462: abundance); Ferrari & Dahms, 2007 (p.64, Rem.: diapause); Hidalgo & Escribano, 2007b (p.501, figs.4, 5, 8); 2008 (p.429: annual life cycle); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Escribano & al., 2009 (p.1083, Table 1, 2, figs.6, 8, 10); C.E. Morales & al., 2010 (p.158, Table 1, 2); Hidalgo & al., 2010 (p.2089, fig.2, 4, Table , cluster analysis2); Tutasi & al., 2011 (p.791, Table 1, 3, fig.10, abundance distribution vs. La Niña event, Rem. p.799, fig.11); Hidalgo & al., 2012 (p.134, Table 2, 3, figs.6, 8, occurrence vs. hydrology); Fierro Gonzalvez, 2014 (p.1, Tab. 3, 5, occurrence, abundance); Bode M. & al., 2014 (p.11, Rem.); Escribano & al., 2014 (p.104, egg production, development vs. temperature); Ruz & al., 2015 (p.200, seasonal abundance vs egg production, hatching success vs. O2); Escribano & al., 2016 (p.1, growth rate) | | | | NZ: | 2 | | |
|
Carte de distribution de Calanus chilensis par zones géographiques
|
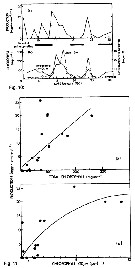
| | | 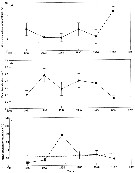 Issued from : R. Escribano & P. Hidalgo inICES J. Mar. Sci., 2000, 57 (6). [p.1872, Fig.3]. Issued from : R. Escribano & P. Hidalgo inICES J. Mar. Sci., 2000, 57 (6). [p.1872, Fig.3].
Annual means of (a) sea surface temperature, (b) female length, anrd (c) abundance of Calanus chilensis from tata obtained on a monthly basis off the Mejillones Peninsula, northern Chile.
The expected abundance of C. chilensis (dashed line) in the lower panel was estimated from the regression of abundance as a function of temperature.
Populations were smallest during the two El Niño events, 1992 and 1997, and largest during the cold year of 1994. The two El Niño events also coincide with the smlallest female sizes observed in 1992 and 1997. |
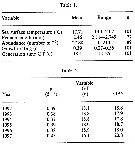 Issued from : R. Escribano & P. Hidalgo inICES J. Mar. Sci., 2000, 57 (6). [p.1870, Table 1 & 2]. Issued from : R. Escribano & P. Hidalgo inICES J. Mar. Sci., 2000, 57 (6). [p.1870, Table 1 & 2].
Table 1: Global means and ranges of sea surface tempetature, female length, and numerical abundance observed from field sampling, and the expected temperature-dependant growth rate and generation time of Calanus chilensis from December 1991 through January 1998 in the coastal zone of the Mejillones Peninsula, northern Chile.
Table 2: Expected annual means of temperature-dependent growth rate (g), generation time (GT) and the number of generations [cohorts] per year (GPY) of Calanus chilensis in the coastal zone of the Mejllones Peninsula, northern Chile.
After Escribano, during the study period (1991-1998), two warm events were reported, in 1991/92 and in 1997/98. The presence of the 1991/92 El Niño was detected in the North Pacific subtropical gyre (Karl & al., 1995) and caused a positive anomaly and deepening of the thermocline during March/April 1992 in coastal waters off Mejillones Peninsula. The 1997-1998 El Niño is considered to be strongest on record (McPhaden, 1999). On the other hand, the sea-surface temperature time-series shows two sharp cold events, in winter 1992 and in winter 1996, whereas 1994 might be considered as a cold year.
Interannually factors other than simply temperature may regulate C. chilensis abundance. |
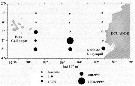 Issued from : P. Tutasi, S. Palma & M. Caceres in Scienc. Mar., 2011, 75 (4). [p.799, Fig.10] Issued from : P. Tutasi, S. Palma & M. Caceres in Scienc. Mar., 2011, 75 (4). [p.799, Fig.10]
Geographic distribution of Calanus chilensis in September and October 2001, associated with the weak La Niña event of 2001. |
 Issued from : W.T. Peterson & D.C. Bellantoni S. Afr. J. Mar. Sci., 1987, 5. [p.418, Figs. 10, 11]; Issued from : W.T. Peterson & D.C. Bellantoni S. Afr. J. Mar. Sci., 1987, 5. [p.418, Figs. 10, 11];
Fig. 10: Temporal changes in (a) egg production of female Calanus chilensis and (b) total chlorophyll at the site off central Chili (off Concepcion).
Fig. 11: Scattergrams of fecundity of C. chilensis against chlorophyll concentration for (a) total chlorophyll and (b) the 20 µm size fraction. |
 Issued from : P.M. Ruz, P. Hidalgo, S. Yàñez, R. Escribano & J.E. Keister in J. Mar. Syst., 2015, 148. [p.208, Fig.6]. Issued from : P.M. Ruz, P. Hidalgo, S. Yàñez, R. Escribano & J.E. Keister in J. Mar. Syst., 2015, 148. [p.208, Fig.6].
Temporal changes of female abundance (Ln number per m3), egg abundance (number per L) and egg production rate in situ samples (EPR) (Ln eggs per female per day) for Calanus chilensis in Mejillones Bay (23°S, 70°25'W). |
 Issued from : S. Ban, C. Burns, J. Castel, Y. Chaudron & al. in Mar. Ecol. Prog. Ser., 1997, 157. [p.289, Table 1]. Issued from : S. Ban, C. Burns, J. Castel, Y. Chaudron & al. in Mar. Ecol. Prog. Ser., 1997, 157. [p.289, Table 1].
Synopsis of feeding/reproduction experiments. A total of 17 diatom and 16 copepod species, representative of a variety of worldwide temperate and subarctic environments (E: estuarine, C: coastal ocean), were screened.
Data for fecundity and hatching success are average values measured at the start and end of the inciubations in a minimum of 3 replicate batches, showing the variation in time of the effects of diatoms on copepod reproduction.
Level of significance of the diatom effect between treatments and controls (e.g. non-diatom diets) in categories I to III was p < 0.01. No.. rank (for comparison with Table 2, p.290).
For the signification of the categories I to IV (fecundity vs. Hatching success vs. Duration of experiment days-1): See Eurytemora affinis, Calanus pacificus, Calanus finmarchicus, Temora stylifera.
List of the 12 marine or estuarine species studied (fresh water excluded. CR): Acartia clausi, A. grani, A. steueri, A. tonsa, Calanus finmarchicus, C. helgolandicus, C. pacificus, Centropages hamatus, C. typicus, Eurytemora affinis, Temora longicornis, T. stylifera. |
 Issued from : S. Ban, C. Burns, J. Castel, Y. Chaudron & al. in Mar. Ecol. Prog. Ser., 1997, 157. [p.290, Table 2]. Issued from : S. Ban, C. Burns, J. Castel, Y. Chaudron & al. in Mar. Ecol. Prog. Ser., 1997, 157. [p.290, Table 2].
Combinations of copepod and non-diatom diets in controls in concentrations ranging from 104 to 105 cells ml-1. Data for fecundity and hatching success are average values measured at the start and end of incubation in a minimum of 3 replicate batches.
N.A: unidentified seston.
No19: In Category II. |
| | | | Loc: | | | SE Pacif., SE Galapagos, G. of Guayaquil, Galapagos-EcuadorPeru, Chile, (N-S, Concepcion, off Valparaiso, off Santiago, St. Vicente Bay, Mejillones region), off Chimbote. | | | | N: | 35 | | | | Lg.: | | | (339) F: 3,6-2,6; M: 3,1-2,7; {F: 2,60-3,60; M: 2,70-3,10} | | | | Rem.: | For Tutasi & al. (2011, p.801) this species could be used as a baseline to indicate future La Niña events in waters of the eastern equatorial Pacific. The species absent on the northern edge of the Equatorial Front and had maxima located on the southern edge and in the central sector of the Equatorial Front (2°N-2°S) in September-October 2001. For the currents in the region see the web site of NOAA Coast Watch (www.aoml.noaa.gov). | | | Dernière mise à jour : 12/12/2020 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2025. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 17 août 2025] © copyright 2005-2025 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
