|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Clausocalanoidea ( Superfamille ) |
|
|
|
Clausocalanidae ( Famille ) |
|
|
|
Pseudocalanus ( Genre ) |
|
|
| |
Pseudocalanus mimus Frost, 1989 (F,M) | |
| | | | | | | Ref.: | | | Frost, 1989 (p.544, figs.F,M); Bucklin & al., 2003 (p.335, tab. 2, fig.3, Biomol.); Aarbakke & al., 2011 (p.1487, Table III, fig.2, genetic analysis); Questel & al., 2016 (p.610, Fig.3, genetic analyse , Table VI, VII, VIII) | 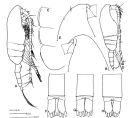 issued from : B.W. Frost in Can. J. Zool., 1989, 67. [Fig.21, p.545]. Female: A, habitus (lateral view); B, anterior portion of cephalosome (lateral view from left side); C, Th1-Th3 (posteroventral margins lateral from left side); . Male: D, habitus (lateral view); E, anterior portion of cephalosome (lateral view from right side); F, posterior portion of urosome; G, idem from Pseudocalanus elongatus; H, idem from Pseudocalanus moultoni. All scale bars = 0.1 mm.
| | | | | Ref. compl.: | | | Shih & Marhue, 1991 (tab.2, 3); Hattori, 1991 (tab.1, Appendix); Gomez-Gutiérrez & Peterson, 1999 (p.637, Table I, II, III, V, VI, figs.4, 5, 6, 7, abundance, egg production); Mackas & al., 2001 (p.685, Tab. 3, fig.3: interannual changes in species composition); Peterson & al., 2002 (p.353); Peterson & al. 2002 (p.381, Table 2, fig.7, interannual abundance); Mackas & Galbraith, 2002 (p.725, tab.2a, 2b, fig.4,5,6); 2002 a (p.423, Table 2); Peterson & Keister, 2003 (p.2499, interannual variability); Mackas & al., 2004 (p.875, Table 2); Mackas & Coyle, 2005 (p.707, fig.7); Mackas & al., 2005 (p.1011, 1021, tab.2); Hopcroft & al., 2005 (p.198, table 2); Mackas & al., 2006 (L22S07, Table 2); Hooff & Peterson, 2006 (p.2610); Mackas & al., 2007 (p.223, climatic change index); Liu & Hopcroft, 2008 (p.928: table II); Peterson, 2009 (p.73, Rem.: p.74, 78, indicator of long-term changes); Galbraith, 2009 (pers. comm.); Hopcroft & al., 2009 (p.9, Table 3); Homma & Yamaguchi, 2010 (p.965, Table 2); Homma & al., 2011 (p.29, Table 2, 3, 4, 5, abundance, feeding pattern: suspension feeders); DFO, 2011 (p.12, Rem.); Bi & al., 2011 (L12607, time series vs decadal oscillation); Matsuno & al., 2011 (p.1349, Table 1, abundance vs years); Matsuno & al., 2012 (Table 1, 2, fig.6); Takahashi M. & al., 2012 (p.393, Table 2, water type index); Irvine & Crawford, 2013 (p.1, Rem.: p.59, anomaly time series); Questel & al., 2013 (p.23, Table 3, interannual abundance & biomass, 2008-2010); Batchelder & al., 2013 (p.34, Rem.: p.42); Hirai & al., 2020 (p.1, Fig. 5: cluster analysis (OTU), spatial distribution). | | | | NZ: | 4 | | |
|
Carte de distribution de Pseudocalanus mimus par zones géographiques
|
| | | 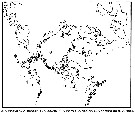 issued from : B.W. Frost in Can. J. Zool., 1989, 67. [p.529, Fig.3]. issued from : B.W. Frost in Can. J. Zool., 1989, 67. [p.529, Fig.3].
Geographical distribution of Pseudocalanus spp. |
 Issued from : W.T. Peterson, J.E. Keister & L.R. Feinberg in Prog. Oceanogr., 2002, 54. [p.391, Fig.7]. Issued from : W.T. Peterson, J.E. Keister & L.R. Feinberg in Prog. Oceanogr., 2002, 54. [p.391, Fig.7].
Abundance (number per cubic meter) of P. mimus at the station NH5 (off Newport, Oregon, at depth 30 m) from May 1996 through September 1999, during El Niño event in 1997-98, the largest of the century.
Nota: The radical changes in community composition observed with the onset of the El Niño were driven in part by the dramatic declines in the abundance of two of the dominant local boreal copepod species, Pseudocalanus mimus and Calanus marshallae (see fig.), from September 1997 through July 1998. Prior to the beginning of the El Niño in 1997, these two boreal species began to increase in abundance in April and by May, the marshallae population had reached 150 ind/m3, and mimus 2500 ind/m3. These values are higher than our 8-year climatology values of 75 and 1500/m3, respectively. Their abundances declines during May/June 1997 downwelling event, rebounded during the brief July/August upwelling period, but then declined to near zero with the onset of downwelling in late-August1997. Their abundances remained persistently low throughout the winter of 1997/98 and most 1998.
Another factor contributing to changeds in community structure during the El Niño period was that C. marshallae and P. mimus were replaced by two warm-water species, C. pacificus (an offshore California Current species) and Paracalanus parvus (a subtropical neritic species). Several other warm-water copepod species occurred commonly during the El Niño period; two of these species, Ctenocalanus vanus and Corycaeus anglicus (see figs.) became common after late August 1997, and were among the most abundant species in the Oregon coastal zone. The boreal coastal copepod species began to recover in July/August of 1998, 11 months after the onset of the El Niño event. C. marshallae numbers began to increase in July and the population appeared to have recovered from the effects of the El Niño by August 1998. Although P. mimus showed signs of a recovery in August 1998, its population did not reach 'normal levels' (based on historical data) until 1999, two years after the advent of the El Niño. Both of the dominant warm-water species, Calanus pacificus and Paracalanus parvus (see figs.), remained abundant through the end of 1998 and persisted in coastal waters (although in low numbers) though mid-summer of 1999. |
| | | | Loc: | | | Japan (NE, off Sanriku); Bering Sea (S, E), Canada Basin (rere); Chukchi Sea, off NW Alaska, S Aleutian Basin, S Aleutian Is., G. of Alaska, British Columbia, Vancouver Island, off Washington coast, Oregon (off Newport) | | | | N: | 33 | | | | Lg.: | | | (122) F: 1,79-1,25; M: 1,29-0,91; {F: 1,25-1,79; M: 0,91-1,29} | | | | Rem.: | Caractéristiques d'après Frost (1989): voir limites des mesures dans la figure au genre Pseudocalanus
Female: partie frontale ogivale en vue latérale; bords postéroventraux de Th1 et Th2 avec un très court processus spiniforme ou absent; rapport des longueurs des segments de P4: B1/B2 < 1,5; rapport des longueurs Prosome/Urosome = 2,10 (1.95-2,24).
Male: rapport des segments longueurs de P4 B1/B2 <1,5; A1: rapport des segments de A1 (8-12)/(20-21) < 2,0; U2-U4 avec sensillum mediodorsal; rapport des longueurs U2/U4 < 1,37; rapport des longueurs Cephalosome/U2 > 5,29 (5,09-5,63); Furca: longueur/largeur > 1,50.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 28/10/2020 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 18 juin 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
{kind=link}
{kind=link}
{kind=link}
