|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Eucalanoidea ( Superfamille ) |
|
|
|
Eucalanidae ( Famille ) |
|
|
|
Eucalanus ( Genre ) |
|
|
| |
Eucalanus californicus Johnson, 1938 (F,M) | |
| | | | | | | Syn.: | Eucalanus giesbrechti Mori, 1937 (1964) (p.22, figs.F, juv.M); Hernandez-Trujillo, 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Hernandez-Trujillo & Esqueda-Escargera, 2002 (in Appendix);
Eucalanus bungii californicus M.W. Johnson, 1938 (p.169, fig.F, Rem.); Davis, 1949 (p.17, figs.F,M); ; Brodsky, 1950 (1967) (p.101, 102, fig.F); Tanaka, 1956 (p.268, figs.F, Rem.); Fleminger, 1967 a (tabl.1); Furuhashi, 1966 a (p.295, vertical distribution vs mixing Oyashio/Kuroshio region, Table 10); Park, 1968 (p.540, Pl. 4, fig.3, Rem.); Morris, 1970 (p.2300); Star & Mullin, 1981 (p.1322, abundance); Van der Spoel & Heyman, 1983 (p.94, fig.58); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Hernandez-Trujillo, 1991 (1993) (tab.I); 1999 (p.284, tab.1); Hernandez-Trujillo & Suarez-Morales, 2002 (p.748, tab.1);
Eucalanus elongatus : Esterly, 1905 (p.131, figs.F,M) | | | | Ref.: | | | Fleminger, 1973 (p.978, 987, 998); Dawson & Knatz, 1980 (p.4, figs.F,M); Gardner & Szabo, 1982 (p.154, figs.F,M); Brodsky & al., 1983 (p.208, figs.F, Rem.F,M); Chihara & Murano, 1997 (p.789, Pl.98,102: F,M); Goetze, 2003 (p.2322 & suiv.) | 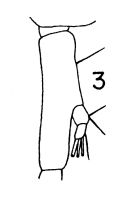 issued from : T. Park in Antarct. Res. Ser. Washington, 1968, 66 (3). [p.539, Pl.4, Fig.3]. As Eucalanus bungii californicus. Female (from tropical-subtropical Central North Pacific): 3, Md (mandibular palp. Nota: The basis of the mandibular palp has a 1 seta.
|
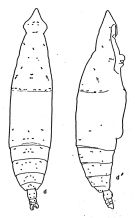 issued from : A. Fleminger in Fishery Bull. natn. Ocean. Atmos. Adm., 1973, 71 (4). [p.989, Fig.13]. Female: d, habitus (dorsal), d', idem (lateral right side).
Dorsal and lateral pattern of integumental organs (black point = sites occuring at 100% frequency, o = 80-99% frequency, x = 10-79% frequency, triangle are sites which are also visible in lateral view but which are assigned to dorsal sets)
|
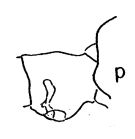 issued from : A. Fleminger in Fishery Bull. natn. Ocean. Atmos. Adm., 1973, 71 (4). [p.968, Fig.1, p (p.969)]. Female: p, genital segment (lateral right side).
|
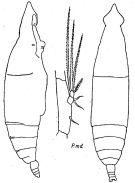 issued from : K.A. Brodsky, N.V. Vyshkvartzeva, M.S. Kos & E.L. Markhaseva in Opred Fauna SSSR, 1983, 135. [p.208, Fig.95]. Female Nota: habitus from Fleminger, 1973 (redrawn); Pmd from Johnson, 1938.
|
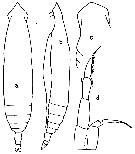 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1905, 2 (4). [p.132, Fig.6]. As .
Female (from San Diego Region): a-b, habitus (dorsal and lateral, respectively).
Male: c, forehead (dorsal); d, P5 (left foot at left of figure).
|
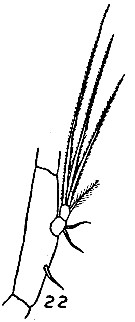 issued from : MW. Johnson in Bull. Scripps Instn Oceanogr., techn. Ser., 4 (6). [p.169, Fig.22]. As Eucalanus bungii californicus. Female (from California): 22, Md (mandibular palp). Nota: Left A1 is the heavier and overreaches the caudal rami by about 5 to 6 segments, wereas the right A1 overreaches the rami by only 3 to 4 segments. 1 single seta on the 2nd mandibular basis of the palp of Md. In the adult Male only a small papilla or scar (after aptophy of the mouth parts), remain to indicate the number of setae that were present in the submatures stages
| | | | | Ref. compl.: | | | Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); Fleminger & Hulsemann, 1973 (p.343, carte); Ohman, 1988 (p.143, Table 1: lipid content); Landry & al., 1994 (p.55, abundance, grazing); Suarez-Morales & Gasca, 1998 a (p.109); Ohman & al., 1998 (p.1709, organic composition, vertical distribution, ETS activity, egg production, dormancy: Table 3); Gomez-Gutiérrez & Peterson, 1999 (p.637, Table I, II, III, VI, figs.4, 5, 6, 7, abundance, egg production); Rebstock, 2001 (tab.2, 4); Peterson & al., 2002 (p.353); Peterson & al. 2002 (p.381, Table 2, interannual abundance); Rebstock, 2002 (p.71, Table 3, 5, 6, Fig..2, climatic variability); Rau & al., 2003 (p.2431, N variations vs climate variability 1950-2000); Peterson & Keister, 2003 (p.2499, interannual variability); Keister & Peterson, 2003 (p.341, Table 1, 2, abundance, cluster species vs hydrological events); Mackas & al., 2004 (p.875, Table 2); Shimode & al., 2005 (p.113 + poster); Lavaniegos & Jiménez-Pérez, 2006 (p.142, tab.2, 3, Rem.); Mackas & al., 2006 (L22S07, Table 2); Ware & McQueen, 2006 (p.28, Table B1, weight ranges); Papastephanou & al., 2006 (p.3078, Table 3); Galbraith, 2009 (pers. comm.); Chiba & al., 2009 (p.1846, Table 1, occurrence vs temperature change); Goetze & Ohman, 2010 (p.2110, Table 1, biogeography); Shimode & al., 2012 a (p.1, life history); Ohman & al., 2012 (p.46, N variations as index of climate variations over 54-year time period); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Kobari & al., 2013 (p.78, Table 2); Hirai & al., 2013 (p.1, Table I, molecular marker); Baumgartner & Tarrant, 2017 (p.387, Table 1, diapause, Rem.); Record & al., 2018 (p.2238, Table 1: diapause); Hirai & al., 2020 (p.1, Fig. 5: cluster analysis (OTU), spatial distribution). | | | | NZ: | 4 | | |
|
Carte de distribution de Eucalanus californicus par zones géographiques
|
| | | | | | 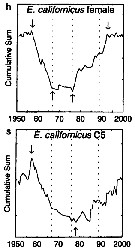 issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 h, s]. issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 h, s].
Climatic regime shifts and decadal-scale variability in calanoid copepod populations off southern California (31°-35°N, 117°-122°W.
Cumulative sums of nonseasonal anomalies from the long-term means of copepod abundance from years 1950 to 2000.
A negative slope indicates a period of below-average anomalies; a positive slope indicates a period of above-average anomalies. Abrupt changes in slope indicate step changes. Step changes are marked with arrows (upward-pointing for increases, downward -pointing for decreases).
The October 1966 cruise (prior to the increase in sampling depth), March 1976 cruise (prior to the 1976-77 climatic regime shift), and October 1988 cruise (prior to the hypothesized 1989 climatic regime shift) are marked with vertical lines. |
 Issued from : S. Shimode, K. Takahashi, Y. Shimizu, T. Nonomura & A. Tsuda in Progress in Oceanography, 2012, 96. [p.11, Fig.10, a]. Issued from : S. Shimode, K. Takahashi, Y. Shimizu, T. Nonomura & A. Tsuda in Progress in Oceanography, 2012, 96. [p.11, Fig.10, a].
Schematic illustration of the most probable life cycle of Eucalanus californicus (a). Ontogenetic vertical migration during the year in the Kuroshio-Oyashio transition area
C = copepodite stage; F = female adult; M = male adult.
Compare with Eucalanus bungii reported in the Oyashio region modified from Tsuda & al. (2004) and Shoden & al. (2005).
Nota:
Gonadal analysis of adult females showed that surface reproduction occurs during the spring in the slope water area, which is a region on the continental slope north of the Kuroshio axis, and within the Kuroshio-Oyashio transition area.
The seasonal ontogenetic vertical migration strategy in the NW Pacific is characterized by shorter periods of surface reproduction and growth, deep copepodite stage 5 dormancy (500-2000 m). This strategy of deep dormancy is considered to be an adaptation to avoid drift caused by a local oceanic feature of deeper westward (> 1000 m) recirculation currents, thus retaining the population within the high primary production zones of the Kuroshio-Oyashio transition area and slope water area. |
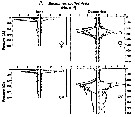 Issued from : M.D. Ohman, A.V. Drits, M.E. Clarke & S. Plourde in Deep-Sea Res. II, , 1998 , 45. [p.1719, Fig. 4 A]. Issued from : M.D. Ohman, A.V. Drits, M.E. Clarke & S. Plourde in Deep-Sea Res. II, , 1998 , 45. [p.1719, Fig. 4 A].
Vertical distribution of copepod in the San Diego Trough in June and December. Adult female and copepodid stage V of Eucalanus californicus.
Dark symbols illustrate nightime distributions, open symbols illustrate daytime distributions, each plotted at the mid-point of the sampling interval. Sampling extended deeper in December than in June.
Sampling out in the San Diego Trough (near 32°50'N, 117°40'W) in 6-14 June 1992 and 15-22 December 1992.
The species is dormant as both adult females and copepodid V's, although winter females respond relatively rapidly to elevated food and temperature conditions; they begin feeding and producing eggs within 2-3 days. |
 Issued from : M.D. Ohman, A.V. Drits, M.E. Clarke & S. Plourde in Deep-Sea Res. II, , 1998 , 45. [p.1730, Table 3]. Issued from : M.D. Ohman, A.V. Drits, M.E. Clarke & S. Plourde in Deep-Sea Res. II, , 1998 , 45. [p.1730, Table 3].
Differential dormancy of co-occuring copepods in the California Current System in the San Diego Trough.
Indices used to differentiate actively growing from dormant animals included developmental stage structure and vertical distribution; activity of aerobic metabolic enzymes (Citrate Synthase and Electron Transfer System complex); investment in depot lipids (wax esters and triacylglycerols); in situ grazing activity from gut fluorescence, and egg production rates. |
| | | | Loc: | | | N Pacif. (Hecate Strait, Oregon (off Newport), California, Santa Monica Basin, W Baja California - Japan), China seas (East China Sea), Japan (Kuroshio-Oyashio trnsition area) | | | | N: | 40 | | | | Lg.: | | | (22) F: 7-5,9; (72) F: 6,4-5,28; (125) F: 6,18; (131) F: 5,9-7; (142) F: 7,5-4,4; M: 4; (866) M: 4,5-5; {F: 4,40-7,50; M: 4,00-5,00} | | | | Rem.: | Entre 30°-50° de latitude nord dans le Pacifique.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 28/10/2020 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 07 avril 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
