|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Eucalanoidea ( Superfamille ) |
|
|
|
Eucalanidae ( Famille ) |
|
|
|
Pareucalanus ( Genre ) |
|
|
| |
Pareucalanus attenuatus (Dana, 1849) (F,M) | |
| | | | | | | Syn.: | Calanus attenuatus (Dana,1849);
Calanus mirabilis Lubbock, 1856;
Calanella mediterranea Claus, 1863 (p.176);
Eucalanus pseudattenuatus Sewell, 1947 (part., p.39, fig. 7A); ? Patel, 1975 (p.659); Zheng & al., 1982 (p.13, figs.F,M); Guangshan & Honglin, 1984 (p.118, tab.); ? Wong & al, 1998 (tab.2); ? Rakhesh & al., 2008 (p.154, Table 5: abundance vs stations);
Eucalanus attenuatus s.s. : Brinton & al., 1986 (p.228, Table 1);
no E. attenuatus : Brodsky, 1962 c (p.113, figs.F); Park, 1968 (p.533: large forms); Bradford, 1970 (p.353); Othman & al., 1990 (p.561, 563, Table 1);
? Eucalanus (Pareucalanus) attenuatus: Seridji & Hafferssas, 2000 (tab.1);
? Eucalanus attenuatus : Brady, 1883 (p., figs.F,M); Giesbrecht, 1892 (p.131, 772, figs.F,M); Giesbrecht & Schmeil, 1898 (p.20, Rem. F,M); Wheeler, 1901 (p.167, fig.F); Thompson & Scott, 1903 (p.233, 242); Cleve, 1904 a (p.189); Esterly, 1905 (p.133, figs.F); Pearson, 1906 (p.7, Rem.); Carl, 1907 (p.16); Wolfenden, 1911 (p.204); Sewell, 1912 (p.353, 357: Rem.); 1914 a (p.200); With, 1915 (p.52, figs. juv.); Sars, 1925 (p.21); Rose, 1925 (p.151); Farran, 1929 (part., p.207, 218); Sewell, 1929 (p.47); Rose, 1929 (p.14); ? Dakin & Colefax, 1933 (p.204); Rose, 1933 a (p.68, figs.F,M); Tanaka, 1935 (p.145, figs.F,M); Farran, 1936 a (p.78); Mori, 1937 (1964) (p.22, figs.F,M, Rem.); ? Dakin & Colefax, 1940 (p.96, figs.F); Wilson, 1942 a (part., p.184); Lysholm & al., 1945 (p.8); Oliveira, 1945 (p.191); Vervoort, 1946 (part., p.95); Brodsky, 1950 (1967) (p.104, figs.F,M); C.B. Wilson, 1950 (part., p.207); Carvalho, 1952 a (p.140, figs.F); Krishnaswamy, 1953 (p.110); King & Hida, 1955 (p.11); Rose, 1956 (p.459); ? Tanaka, 1956 (p.267, figs.M, Rem.); Chiba & al., 1957 (p.306); 1957 a (p.11); Yamazi, 1958 (p.147, Rem.); Marques, 1958 (p.206); Deevey, 1960 (p.5, Table II, annual abundance); Heinrich, 1961 b (p.93); Grice & Hart, 1962 (p.287, table 3); Grice, 1962 (p.180, figs.F, Rem.); Fagetti, 1962 (p.9); Cervigon, 1962 (p.181, tables: abundance distribution); Fish, 1962 (p.10); Ganapati & Shanthakumari, 1962 (p.7, 15); ? V.N. Greze, 1963 a (tabl.2); Vervoort, 1963 b (p.90, fig.M, Rem.); ? Gaudy, 1963 (p.20); ? Björnberg, 1963 (p.19, Rem.); Unterüberbacher, 1964 (? part., p.15, Rem) ; De Decker, 1964 (? part., p.15, 22, 30); De Decker & Mombeck, 1964 (p.12); Chen & Zhang, 1965 (p.33, figs.F,M); Mullin, 1966 (p.546, Table I, III, diet); Mazza, 1966 (p.69); 1967 (p.65, figs.juv., F,M); Furuhashi, 1966 a (p.295, vertical distribution in Kuroshio region, Table 10); Saraswathy, 1966 (1967) (p.75); Neto & Paiva, 1966 (p.18, Table III, annual cycle); Fleminger, 1967 a (tabl.1); Grice & Hulsemann, 1967 (p.13); ? Ehrhardt, 1967 (p.736, geographic distribution, Rem.); Park, 1968 (p.533, figs.F: small form); ? De Decker, 1968 (p.45); Marques, 1968 a (p.4); Timonin, 1971 (p.281, trophic group); Evans, 1968 (p.13); Vinogradov, 1968 (1970) (p.78, 266); Delalo, 1968 (p.137); Park, 1968 (p.533, figs.: small forms); Bradford, 1970 a (p.353, figs.F,M); Park, 1970 (p.474); Morris, 1970 (p.2300); Corral Estrada, 1970 (p.73, figs.F,M); Dowidar & El-Maghraby, 1970 (p.268); Shih & al., 1971 (p.39); Gueredrat, 1971 (p.300, fig.3, 10, Table 1, 2); Deevey, 1971 (p.224); Binet & al., 1972 (p.68); Roe, 1972 (tabl.1, tabl.2); 1972 a (p.324); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night, Table II: %); Nival & al., 1972 (p.63, respiration); ? Roe, 1972 (p.277, tabl.1, tabl.2); Björnberg, 1972 (p.13, figs.N); ? 1973 (p.296, 386 Rem.); ? Guglielmo, 1973 (p.399); Heinrich, 1973 (p.95, fig.3); Marques, 1973 (p.234, fig.M juv.); Geletin, 1973 (p.1783, 1786, figs., Rem.); Fleminger, 1973 (p.978, 995, 998, figs.F,M, fig.47, Rem.); ? Harding, 1974 (p.141, Table 2, gut contents); Yamazi, 1974 (p.41); Marques, 1974 (p.12); Vives & al., 1975 (p.35, tab.II); Fleminger, 1975 (p.395); Patel, 1975 (p.659); Arashkevich, 1975 (p.351, digestion time); ? Ikeda, 1976 (p.51, respiration rate); Geletin, 1976 (p.76, 82, 91); Marques, 1976 (p.987); Greenwood, 1976 (p.10, figs.F); Timonin, 1976 (p.79, vertical distribution); Tranter, 1977 (p.596); Madhu Pratap & al., 1977 (p.138, Table 3: abundance vs. stations); Timonin & Voronina, 1977 (p.285, fig.3); Frontier, 1977 a (p.14); Dessier, 1979 (p.204); Goswami & Goswami, 1979 (p.103, figs.); Svetlichnyi, 1980 (p.28, Table 1 as Fucalanus attenuatus, passive submersion); Vaissière & Séguin, 1980 (p.23, tab.2); Sreekumaran Nair & al., 1981 (p.493); Madhupratap & al., 1981 (p.262, Table 2: abundance vs. thermocline); Madhupratap & al., 1981 (p.266, fig.2e: abundance vs. geographic transect); Marques, 1982 (p.752); Vives, 1982 (p.290); Kovalev & Shmeleva, 1982 (p.82); Zheng & al., 1982 (p.11, figs.F,M); Brodsky & al., 1983 (p.208, figs.F,M, Rem.); Van der Spoel & Heyman, 1983 (p.42, fig.54); Gaudy & Boucker, 1983 (p.37, Table 1, 3, fig.1, 2, Rem.: metabolism); Dessier, 1983 (p.89, Tableau 1, 2, Rem., %); Tremblay & Anderson, 1984 (p.5: Rem.); Stephen, 1984 (p.161, 169, Distribution vs thermocline & geographic); Guangshan & Honglin, 1984 (p.118, tab.); Sameoto, 1984 (p.767, vertical migration); De Decker, 1984 (? part., p.315, 346: carte); Sazhina, 1985 (p.32, figs.N); Longhurst, 1985 (tab.2); Kimmerer & al., 1985 (p.428); Price & Paffenhöfer, 1985 (p.115, feeding); Regner, 1985 (p.11, Rem.: p.23); Baars & Oosterhuis, 1985 (p.71, Table 4: gut passage time); Petipa & Borichenko, 1985 (tab.1); Baessa de Aguiar, 1986 (1989) (p.59, Rem.); Madhupratap & Haridas, 1986 (p.105, tab.1); Renon, 1987 (tab.2); Cowles & al., 1987 (p.653, Table 4, egg production); Valentin & al., 1987 (p.1202); Lozano Soldevilla & al., 1988 (p.57); Jimenez-Perez & Lara-Lara, 1988; Dessier, 1988 (tabl.1); Hernandez-Trujillo, 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); ? Suarez & al., 1990 (tab.2); Timonin, 1990 (p.479); Schulz, 1990 (p.188); Hirakawa & al., 1990 (tab.3); Scotto di Carlo & al., 1991 (p.271); Huys & Boxshall, 1991 (p.60, fig.M); Yoo, 1991 (tab.1); Baessa De Aguiar, 1991 (1993) (p.106); Hernandez-Trujillo, 1991 (1993) (tab.I); Samba Diouf, 1991 (p.103, tableau 1); Ayukai & Hattori, 1992 (p.163, Table 5, fecal pellet production rate); Seguin & al., 1993 (p.23); Kim & al., 1993 (p.269); Webber & Roff, 1995 (tab.1); Webber & al., 1996 (tab.1); Pakhomov & McQuaid, 1996 (p.271, abundance, distrbution, seabirds); Shih & Young, 1995 (p.69); Madhupratap & al., 1996 (p.866); Padmavati & Goswami, 1996 a (p.85, fig.3, Table 4, vertical distribution); Kotani & al., 1996 (tab.2); Go & al., 1997 (tab.1); Park & Choi, 1997 (Appendix); ? Chihara & Murano, 1997 (p.788, Pl.99,102: F,M); Noda & al., 1998 (p.55, Table 3, occurrence); Padmavati & al., 1998 (p.349); ? Gilabert & Moreno, 1998 (tab.1, 2); Mauchline, 1998 (tab.8, 23, 30, 64); Hernandez-Trujillo, 1999 (p.284, tab.1); Dolganova & al., 1999 (p.13, tab.1); Onishchik, 1999 (p.76, figs. 2, 5); Fernandez-Alamo & al., 2000 (p.1139, Appendix); Escribano & Hidalgo, 2000 (p.283, tab.2); Hidalgo & Escribano, 2001 (p.158, fig.5); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); ? Moraitou-Apostolopoulou & al., 2000 (tab.I); Madhupratap & al., 2001 (p. 1345, vertical distribution vs. O2, figs.4, 5: clusters); Rebstock, 2001 (tab.2); Holmes, 2001 (p.40); Nakata & al., 2001 (p.335, tab.4, 5, 6, 7); Hidalgo & Escribano, 2001 (p.159, tab.2); Lo & al., 2001 (1139, tab.I); ? Sameoto & al., 2002 (p.12); ? Beaugrand & al., 2002 (p.179, figs.5, 6); Rebstock, 2002 (p.71, Table 3, climatic variability); Hernandez-Trujillo & Esqueda- Escargera, 2002 (in Appendix); Osore & al., 2003 (p.69);; Vukanic, 2003 (139, tab.1); Eazai & al., 2004 (p.489, tab.2); ? G. Harding, 2004 (p.40, figs.F,M); Shimode & al., 2005 (p.113 + poster); ? Berasategui & al., 2005 (p.485, tab.1); Escribano, 2006 (p.20, Table 1); ? Koppelmann & Weikert, 2007 (p.266: tab.3); no Isari & al., 2006 (p.241, tab.II); Ferrari & Dahms, 2007 (p.70, 71, Rem.); Zakaria & al., 2007 (p.52, Table 1, vs Salinity); Vives & Shmeleva, 2007 (p.873, figs.F,M, Rem.); Rakhesh & al., 2008 (p.154, Table 5: abundance vs stations); Morales-Ramirez & Suarez-Morales, 2008 (p.519); Fernandes, 2008 (p.465, Tabl.2); Wishner & al., 2008 (p.163, Table 2, fig.8, oxycline); Pagano, 2009 (p.116); Licandro & Icardi, 2009 (p.17, Table 4); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Fazeli & al., 2010 (p.153, Table 1); Shanthi & Ramanibai, 2011 (p.132, Table 1); Maiphae & Sa-ardrit, 2011 (p.641, Table 2); Dorgham & al., 2012 (p.473, Table 4: abundance vs season); Kobari & al., 2013 (p.78, Table 2); Williams R., 2013 (p.71, figs., Appendix A, C; C & N composition, trophic level); Aboul Ezz & al., 2014 (p.283, Rem.: p.284); Zaafa & al., 2014 (p.67, Table I, occurrence); Acha & al., 2020 (p.p.1, Table 3: occurrence % vs ecoregions).
Subeucalanus attenuatus : Hwang & al., 2003 (p.193, tab.2); Tseng & al., 2009 (p.327, fig.5, feeding);
Paracalanus attenuatus : Tseng & al., 2008 (p.402, Table 2);
Euchaeta attenuatus : Huntley & Lopez, 1992 (p.201, Table B1, growth rate, temperature-dependent production [lapsus calami]); Paraeucalanus attenuatus : Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Tutasi & al., 2011 (p.791, Table 1, 3, figs.9, 11, abundance distribution vs La Niña event); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters);
Eucalanus attenuates : Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution);
Eucalanus attenuatus s.l. : Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution);
| | | | Ref.: | | | Fleminger, 1973 (p.990, figs.F); Geletin, 1976 (p.83, 91); Bradford-Grieve, 1994 (p.80); Conway & al., 2003 (p.164, figs.F,M, Rem.); Goetze, 2003 (p.2322 & suiv.); Boxshall & Halsey, 2004 (p.113: figs.F); Mulyadi, 2004 (p.107, figs.F,M, Rem.); Prusova, 2007 (p.64, tab.1: integumental perforation pattern); Phukham, 2008 (p.123, figs.F, M); Goetze & Ohman, 2010 (p.2110, Table 1, A1, Fig.11, molecular study, biogeography); Andronov, 2014 (p.36, fig.: A2) | 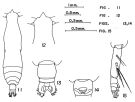 issued from : T. Park in Antarct. Res. Ser. Washington, 1968, 66 (3). [p.531, Pl.1, Figs.11-15]. As Eucalanus attenuatus small form. Doubtful. Female from Central North Pacific: 30°-31°N, 155°-154°W): 11, habitus (dorsal); 12, forehead (dorsal); 13, urosome (dorsal); 14, genital segment ( (ventral); 15, Md (mandibular palp). Nota: Proportional lengths of prosome to urosome are about 7.2:1; The 2 free urosomal segments and the caudal furca are 49:16:35. The genital segment is wider than long (55:45). The mandibular palp has 2 setae on the basis, 4 setae on the 2nd endopodal segment, and 6 setae on the exopod.
|
 issued from : A. Fleminger in Fishery Bull. natn. Ocean. Atmos. Adm., 1973, 71 (4). [p.968, Fig.1, q (p.969)]. Female (from Pacific: 13°23'S, 126°W): q, genital segment (lateral right side).
|
 issued from : A. Fleminger in Fishery Bull. natn. Ocean. Atmos. Adm., 1973, 71 (4). [p.994, Fig.18, a]. As Eucalanus attenuatus. Female (from 00°02'N, 166°59'W): a, forehead (lateral right side).
|
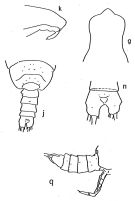 issued from : A. Fleminger in Fishery Bull. natn. Ocean. Atmos. Adm., 1973, 71 (4). [p.994, Fig.18]. As Eucalanus attenuatus. Male: g, forehead (dorsal); j, 4th and 5th toracic segments and uropode (dorsal); k, forehead (lateral right side); n, anal segment and fused furcal rami (dorsal); q, urosome and P5 (lateral right side). g, j, q from specimens from 00°02'N, 166°59'W; n, specimen from 39°18'S, 119°51'E. Nota: left P5 with 4 segments, right P5 about half as long and 3 segments.
|
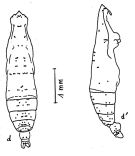 issued from : A. Fleminger in Fishery Bull. natn. Ocean. Atmos. Adm., 1973, 71 (4). [p.991, Fig.15 Ad, Bd]. As Eucalanus attenuatus. Female: a, habitus (dorsal); a', idem (lateral right side). Dots represent sites occuring at a frequency of 100% in the pooled sample of the species, open circles are sites appearing in 80 to 99%, crosses are sites occupied in from 10 to 79%. Open triangles are sites also visible in lateral view but assigned to tergal sets.
|
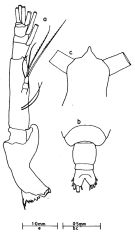 issued from : J.G. Greenwood in Proc. R. Soc. Qd, 1976, 87. [p.13, Fig.4, a-c]. As Eucalanus attenuatus. Female (from Moreton Bay, E Australia): a, Md; b, posterior metasome and urosome (dorsal); c, forehead (dorsal).
|
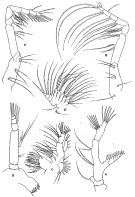 issued from : R.B.S. Sewell in The John Murray Expedition, 1933-34, Scientific Reports, VIII (1), 1947. [p.41, Fig.8]. As Eucalanus pseudattenuatus. Female (from Arabian Sea): A, A2; B, Md; C, Mx1; D, Mx2; F, Mxp. Copepodid stage V: E, Mxp.
|
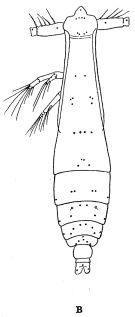 issued from : R.B.S. Sewell in The John Murray Expedition, 1933-34, Scientific Reports, VIII (1), 1947. [p.39, Fig.7, B]. As Eucalanus pseudattenuatus. Female (fromArabian Sea): B, habitus (dorsal). Nota:The total length of the females ranges from 3.00 to 3.383 mm; male: 3.067.
|
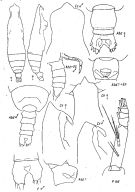 issued from : K.A. Brodsky, N.V. Vyshkvartzeva, M.S. Kos & E.L. Markhaseva in Opred Fauna SSSR, 1983, 135. [p.209, Fig.96]. Female & Male; Nota: Ce, Abd+Gn, P md, from Park, 1968; other figures from Fleminger, 1973. (redrawn).
|
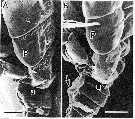 issued from : S. Ohtsuka & R. Huys in Hydrobiologia, 2001, 453/454. [p.450, Fig.5, A-B]. As Eucalanus attenuatus. Female (from Japan): A, pedigers 2-5 (p) and urosome (u), lateral. Male: B, pedigers 3-5 (p) and urosome (u), lateral (P5 arrowed). Nota: In female, the dorsal and lateral sides of the last three prosomites (pedigers 2-5) are covered with minute spinules, which may function in helping the male to grasp the female, as in the Cala,idae (Tsuda & Miller, 1998). These spinules are completely lacking in the male. The Mxp is relatively shorter in the male than in the female, and the plumose setae on the basis and endopod are much better developed in the male. The inwardly directed plumose on the basis and endopod of the male Mxp may function in grasping, whereas outwardly directed setae on the 5th and 6th endopodal segments may play a restricted role in generating a scanning current, togeher with the other appendages of the head.
|
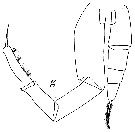 issued from : W. Vervoort in Atlantide Report., 1963, 7. [p.197, Fig.3, g]. As Eucalanus attenuatus. Male (from off Ghana): g, P5 (x220).
|
 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.12, Fig.4-1]. As Eucalanus attenuatus. Doubtful. Female: a, habitus (dorsal); b, urosome (ventral); c, forehead (lateral); d, urosome (lateral, right side); e, A2; f, Md (mandibular palp); g, Md (endopod of mandibular palp); h, Mx1; i, P1; j, P2. Scale bar in mm. Nota: length: 5.78 mm.
|
 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.13, Fig.4-2]. As Eucalanus attenuatus. Doubtful. Male: k, habitus (dorsal); l, forehead (dorsal); m, idem (dorsal, another specimen); n, idem (lateral); o, urosome (dorsal); p, A2; q, Md (mandibular palp); r, endopod of mandibular palp of Md; s, Mx1. Scale bar in mm. Nota: length; 4.50 mm.
|
 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.14, Fig.5-1]. As Eucalanus pseudattenuatus. Dounbtul. Female: a, habitus (dorsal); b, forehead (lateral); c-d, urosome (lateral and ventral, respectively); e, A2; f, Md; g, endopod of Md; h, Mx1; i, P1; j, P2. Scale bar in mm. Nota: length : 4,14 mm
|
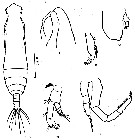 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.15, Fig.5-2]. As Eucalanus pseudattenuatus. Doubtful. Male: k, habitus (dorsal); l, forehead (lateral); m, A2; n, Md (mandibular palp); o, endopod of Md; p, P5. Scale bar in mm. Nota: length: 2.97-3.20 mm.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.8, Figs.1-6]. As Eucalanus attenuatus. Female: 3, habitus (dorsal); 5, Md; 6, Mx1. Nota: Forehead triangular, aand sometimes indented on each side.1 segment between the anal and genital segments which is longer than its broad. 1st segment of endopodite of A2 is 4 times as long as its broad, and longer than the 2nd segment. Inner margin of the 2nd segment of basipodite of Md divided into approximately equal portions by the insertion of the endopodite; 2nd segment of endopodite with 4 setae on its apex. Mx1 with 2nd lobe present; 2nd segment of basipodite with 5 inner marginal setae. Male: 1-2, habitus (lateral and dorsal, respectively); 4, P5.
|
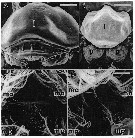 issued from : S. Ohtsuka & R. Huys in Hydrobiologia, 2001, 453/454. [p.459, Fig.1]. As Eucalanus attenuatus. Female from Japan): A, labrum; C, dorsal wall of filter basket between maxillae (ma) and maxillipeds (mp). Male (from Japan): B, labrum (1) and paragnath (p); D, dorsal wall of filter basket between maxillae (ma) and maxillipeds (mp). Scale bars: 0.1 mm. Nota: On the ventral integumental wall between Mx2 and Mxp of female, fine spinules and setules are present. These are lacking in the non feeding male. Probably these structures reduce leakage of particles captured within the 'filter basket'. The posterior margin of the labrum in the female bears short, stout spinules directed toward the oesophagus, whereas that of the non-feeding male is naked. This presumably also plays a role in reducing leakage of food particles during chewing with the mandibular gnathobases. Example of seexual trophic dimorphism.
|
 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.179, Pl.2, Figs.10-11]. As Eucalanus attenuatus. With doubt. Female (from equatorial Pacific): 10, 2nd basal segment and endopod of Md; 11, Mx1.
|
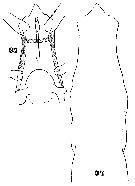 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 35, Figs.34, 37]. As Eucalanus attenuatus. With doubt. See in remarks. Female: 34, head (dorsal); 37, forehead (ventral).
|
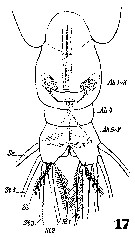 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 35, Fig.17]. As Eucalanus attenuatus. With doubt. See in remarks. Female: 17, urosome (ventral).
|
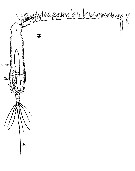 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 35, Fig.3]. As Eucalanus attenuatus. With doubt. See in remarks. Male: 3, habitus (dorsal).
|
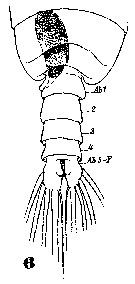 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 35, Fig.6]. As Eucalanus attenuatus. With doubt. See in remarks. Male: 6, distal thoracic segments and urosome (dorsal). F = caudal ramus.
|
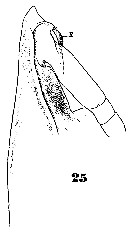 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 35, Fig.25]. As Eucalanus attenuatus. With doubt. See in remarks. Female: 25, forehead (lateral).
|
 issued from : R. Huys & G.A. Boxshall in Copepod Evolution. The Ray Society, 1991, 159. [p.60, Fig.2.2.7, D]. As Eucalanus attenuatus. Male (from John Murray Expedition, 1933-34): D, right A1.
|
 Issued from : G.S. Brady in Rep. Scient. Results Voy. Challenger, Zool., 1883, 8 (23). [Pl. VI]. As Eucalanus attenuatus. Doubtful. Female: 1, habitus (lateral); 2, forehead and rostrum (ventral); 3, rostrum; 4, A2; 5, Md; 6, Mx2. Male: 7, Mxp; 8, one of the swimming leg.
|
 Issued from : G.S. Brady in Rep. Scient. Results Voy. Challenger, Zool., 1883, 8 (23). [Pl. II, Figs.8-10]. As Eucalanus attenuatus. With doubt. Female: 8, Mx1; 9, Mxp. Male: 10, P5.
|
 issued from : G. Harding in Key to the adullt pelagic calanoid copepods found over the continental shelf of the Canadian Atlantic coast. Bedford Inst. Oceanogr., Dartmouth, Nova Scotia, 2004. [p.40]. As Eucalanus attenuatus. With doubt. Female & Male. Nota: In both sexes: Posterior border of metasome rounded. Exopod of A2 just reaching the distal margin of the endopodal segment 1.. Setae formula of Md 2: 0: 4.
|
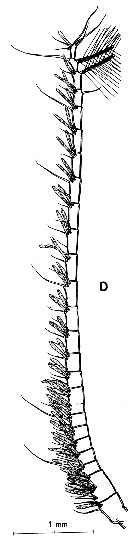
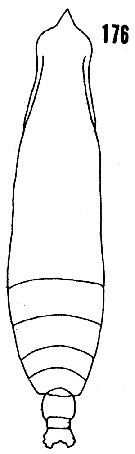 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1967, 1, Crustacea, 1: Copepoda. [p.35, Fig.176]. As Eucalanus attenuatus. Female (from Florida Current): 176, habitus (dorsal). Nota: One segment between the genital and anal segments.
|
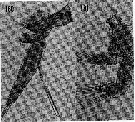 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1967, 1, Crustacea, 1: Copepoda. [p.36, Figs.180-181]. As Eucalanus attenuatus. Female: 180, Md. Nota: Inner margin of basis of Md divided into 2 nearly equal portions by insertion of endopodite. Male: 181, A2. Nota: Exopodite of A2 just reaching distal margin of first segment of endopodite.
|
 issued from : N. Phukham in Species diversity of calanoid copepods in Thai waters, Andaman Sea (Master of Science, Univ. Bangkok). 2008. [p.205, Fig.79]. Female (from W Malay Peninsula): a, habitus (dorsal); b, forehead (lateral); c, Md (mandibular palp); d, Md (cutting edge of gnathobase); e, urosome (dorsal). Male: f, habitus (dorsal); g, urosome (dorsal); h, P5. Body length after drawings: F = 4.159 mm; M = 3.060 mm.
|
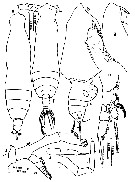 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.107, Fig.59]. Female (from Indonesian Seas): a, habitus (dorsal); b, forehead (lateral); c, urosome (lateral right side); d, A2; e, P1. Male: f, habitus (dorsal); g, forehead (lateral); h, right P5; i, left P5. Nota Female: Prosome 8 times longer than urosome. Forehead apex acute and not curved ventrally. A1 23-segmented, segments 1 and 2, 8 and 9 fused, reaching caudal rami by the last 6-7 segments. A2 endopod segment 1 4 times length of wide, and 1/3 times length of endopod segment 2. Basis of Md with 2 inner setae. Urosome segment 1 (fused somites 1 and 2) longer than wide, longer than anal segment (fused somites 4 and 5) and caudal rami combined. Nota Male: Forehead apex broadly rounded in dorsal and lateral views. Endopod of Md places near distal end of basis. Caudal rami asymmetrical. P1-P4 as in female. P5 left leg 4-segmented, twice longer than right leg; right leg 3-segmented.
|
 Issued from : V.N. Andronov in Russian Acad. Sci. P.P. Shirshov Inst. Oceanol. Atlantic Branch, Kaliningrad, 2014. [p.36, Fig.9, 13]. Pareucalanus attenuatus after Sewell, 1947. 13, exopod of A2.
| | | | | Ref. compl.: | | | Madhupratap & Haridas, 1990 (p.305, fig.6: vertical distribution night/day; fig.7: cluster); ? Siokou-Frangou, 1997 (tab.1); ? Lapernat & Razouls, 2001 (p.123, tab.1); Hsieh & al. 2004 (p.397, tab. 1); Lan & al., 2004 (p.332, tab.1); Gallienne & al., 2004 (p.5, tab.3); Lo & al., 2004 (p.89, tab.1); Kazmi, 2004 (p.229); Prusova & Smith, 2005 (p.76); ? Berasategui & al., 2006 (p.485: fig.2); Zuo & al., 2006 (p.159, tab.1, abundance, fig.8: stations group); Lavaniegos & Jiménez-Pérez, 2006 (p.151, tab.2, 3, Rem.); Hwang & al., 2006 (p.943, tabl.I); Mackas & al., 2006 (L22S07, Table 2); Rakhesh & al., 2006 (p.93, Table 2, spatial distribution); Hwang & al., 2007 (p.24); Dur & al., 2007 (p.197, Table IV); McKinnon & al., 2008 (p.844: Tab.1, p.848: Tab. IV); Tseng L.-C. & al., 2008 (p.153, Table 2, occurrence vs geographic distribution); C.-Y. Lee & al., 2009 (p.151, Tab.2); Escribano & al., 2009 (p.1083, Table 1, figs.6, 7, 10, as Paraeucalanus); C.E. Morales & al., 2010 (p.158, Table 1); Cornils & al., 2010 (p.2076, Table 3); Xu & Gao, 2011 (p.514, figs.3, 4, Table 2: optimal salinity); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); ? Mazzocchi & Di Capua, 2010 (p.425); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); W.-B. Chang & al., 2010 (p.735, Table 2, abundance); Nowaczyk & al;, 2011 (p.2159, Table 2); Hsiao S.H. & al., 2011 (p.475, Appendix I: as Paraeucalanus, lapsus calami); Guo & al., 2011 (p.567, table 2, indicator); Uysal & Shmeleva, 2012 (p.909, Table I); Mulyadi & Rumengan, 2012 (p.202, Rem.: p.204); ? Miloslavic & al., 2012 (p.165, Table 2, transect distribution); ? Belmonte & al., 2013 (p.222, Table 2, abundance vs stations); Tseng & al., 2012 (p.621, Table 3: abundance); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Tachibana & al., 2013 (p.545, Table 1, Rem.: s.l., seasonal change 2006-2008); Tseng & al., 2013 (p.507, seasonal abundance); Tseng & al., 2013 a (p.1, Table 3, 5, abundance); Rakhesh & al., 2013 (p.7, Table 1, abundance vs stations); Jagadeesan & al., 2013 (p.27, Table 3, 5, 6, fig.11, seasonal variation); Anjusha & al., 2013 (p.40, Table 3, abundance & feeding behavior); Lidvanov & al., 2013 (p.290, Table 2, % composition); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Zakaria & al., 2016 (p.1, Table 1); Jackson & Smith, 2016 (p.305, figs. 3, 4, 5, vertical abundance vs day/night cycle); El Arraj & al., 2017 (p.272, table 2, spatial distribution); ? Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); ? Belmonte, 2018 (p.273, Table I: Italian zones); ? Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence). | | | | NZ: | 9 + 3 douteuses | | |
|
Carte de distribution de Pareucalanus attenuatus par zones géographiques
|
| | | | | | | | | | | | 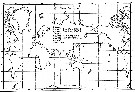 issued from : A. Fleminger in Fishery Bull., 1973, 71 (4). [p.993, Fig.17]. As Eucalanus attenuatus. issued from : A. Fleminger in Fishery Bull., 1973, 71 (4). [p.993, Fig.17]. As Eucalanus attenuatus.
Geographical localities of the species in the attenuatus group. All identifications except those of Park (1968) and Bradford (1970) verified by examination of integumental organs. |
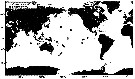 Issued from : E. Goetze & M.D. Ohman in Deep-Sea Res. II, 2010, 57. [p.2123, Fig.11]. Issued from : E. Goetze & M.D. Ohman in Deep-Sea Res. II, 2010, 57. [p.2123, Fig.11].
Biogeographic distribution of Pareucalanus attenuatus, a subtropical-tropical species in the Indian and Pacific Oceans.
Solid circles mark sampling locations of specimens whose species identity was verified by DNA sequencing of 165 rRNA.
Fleminger (1973) records of this species are plotted as open square. |
 Issued from : P. Tutasi, S. Palma & M. Caceres in Scienc. Mar., 2011, 75 (4). [p.799, Fig.9 a] Issued from : P. Tutasi, S. Palma & M. Caceres in Scienc. Mar., 2011, 75 (4). [p.799, Fig.9 a]
Geographic distribution of Paraeucalanus attenuatus in September and October 2001, associated with the weak La Niña event of 2001. |
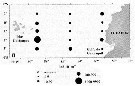
 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 61- Pareucalanus attenuatus.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
 Issued from : Y.-Q. Chen in CalCOFI Rep., 1986, XXVII. [Fig.4, p.212]. As Eucalanus attenuatuss. l. (= Pareucalanus attenuatus s.l.) : Vertical abundance of females, males and late copepodites. Issued from : Y.-Q. Chen in CalCOFI Rep., 1986, XXVII. [Fig.4, p.212]. As Eucalanus attenuatuss. l. (= Pareucalanus attenuatus s.l.) : Vertical abundance of females, males and late copepodites.
Nota: This category includes E. attenuatus s.s. and E. sewelli s.s. Percentage in the samples from the cruise: 2.6 (day: 2.9, night: 2.2). Distributed mainly between 25 and 100 m. Their vertical distribution was mainly confined to depths above 100 m at stations 14-16 (see chart 1 in Subeucalanus subtenuis), they appeared only in the surface between 18°N and 20°N. The distribution of males and females was similar, but the females were more numerous.. At the mouth of the Gulf of California they were found only between 25 and 75 m at station 10.
The maximum concentration was 30,700 individuals /1000 m3.The species were not observed in samples from the oxygen-deficient layer or from stations near Mexico.
(see hydrologic conditions in Subeucalanus subtenuis figs.12, 13) |
| | | | Loc: | | | South Africa (S, E & ? W), ? Atlant., ? G. of Guinea, Morocco-Mauritania, ? Ibero-moroccan Bay, ? Off W Tangier, ? Medit., ? Adriatic Sea, ? Alexandria, W Egyptian coast, off Sharm El-Sheikh, Taba, Red Sea, Gulf of Oman, G. of Aden, Arabian Sea, Arabian Gulf, Madagascar (Nosy Bé), Rodrigues Is. - Seychelles, Mascarene Basin, Kavarattri & Agatti Is.equatorial Indian Ocean, India (Goa - Gujarat, Lawson's Bay, Madras, Gulf of Mannar, Palk Bay, Godavari region, Kakinada Bay), Bay of Bengal, Andaman Sea, W Malay Peninsula (Andaman Sea), Straits of Malacca, Indonesia-Malaysia, Flores Sea, SW Celebes, Manado Bay, N Banda Sea (Ambon Bay), W New-Guinea, Australia (North West Cape), Viet-Nam (Cauda Bay), China Seas (Yellow Sea, East China Sea, South China Sea), Taiwan Strait, Taiwan (E, S, W, SW, NW, N, Mienhua Canyon, NE), Okinawa, S Korea, Korea Strait, Japan Sea, off Japan, Japan (Onagawa, Tokyo Bay, Toyama Bay), off S Shikoku Is., Kuchinoerabu Is., Kuroshio region, Pacif. (equatorial & subtropical), off Hawaii N & S, Carolines Is. - Marshall Is., California, W Baja California, Gulf of California, G. of Tehuantepec, off W Guatemala, W Costa Rica, Clipperton Is., Australia (Shark Bay, G. of Carpentaria, ? New South Wales, Great Barrier), New Caledonia, ? New Zealand, S Pacific (NPFZ), Galapagosd-Ecuador, Peru, Chile (? N, S, Concepcion) | | | | N: | 318 ? (any confusion with other forms) | | | | Lg.: | | | (11) F: 3,383-3; (28) F: 5,6-3,55; M: 5,8-2,75; (72) F: 4,7-4,51; (79) F: 4,65-3,63; M: 3,5-2,93; (91) F: 4-5,5; M: ± 3,5; (101) F: 4,28-3,06; M: 3,06-2,92; (104) F: 5,9; (125) F: 4,76; M: 4,54-4,4; F: 6,19-5,72; (131) F: 3,63-4,65; M: 2,93-3,50; ? (199) F: 6,4-4,33; M: 5,6-3,04; (290) F: 5,1-6; M: 4,25-4,35; ? (432) F: 7,3-5; (530) F: 4,3-4,1; ? (785) F: 4,35-4,08; (795) F: 4; M: 3; (991) F: 3,55-7; M: 2,95-6,04; (1023) F: 5,78; M: 4,50; F: 3,9-3,94; (1122) F: 3,9-3,94; {F: 3,00-7,30; M: 2,75-6,04} | | | | Rem.: | L'analyse des structures tégumentaires conduit Fleminger (1973) à restreindre la diagnose de cette forme, et de ce fait sa répartition géographique.
Cette espèce a été confondue avec Eucalanus parki de plus grande dimension et E. sewelli dont elle ne diffère que légèrement par la forme de la tête (cf. in Bradford-Grieve, 1994: fig.44: B).
Giesbrecht (1892) la signale au nord des Iles du Cap Vert, Wolfenden (1911) près des Açores, Fish (1962, p.3, 10), Sander & Moore (1978, p.232) et Deevey (1960, p.16) la signalent (sans précision morphologique) à l'île Barbade et à l'extérieur de la Baie Delaware en s'appuyant sur le travail de Wilson (1932 a, p.32) off Woods Hole, dont la synonymie reste à préciser. Elle est également signalée dans la Mer des Antilles et le Golfe du Mexique par Park (1970), à la Jamaïque par Webber & Roff (1995), Hopcroft & al., 1998 (tab.2), le courant de Floride par Owre (1962, p.491) et Owre & Foyo (1967, p.35, Rem.), off Cap Cod (in Harding, 1974 (p.141, tab.2); off Bermuda par Deevey, (1971); dans la Baie Delaware par Deevey (1960); En Angola (Baia Farta) par neto & Paiva (1966); Vervoort (1963 b) dans le Golfe de Guinée, Bowman (1971 b, p.5), Binet & al., 1972 (p.68), dans le golfe du Maine par Bigelow (1926, p.228), off Nova Scotia par Sameoto & al., 2002 (p.12) et déjà par Sars (1925, p.21) aux Açores et Pearson (1906, p.7) et Farran (1908b) à louest de lIrlande, par Seguin (1966, p.37) à Dakar; aux Canaries par Roe (1972) et Corral Estrada & Pereiro Muñoz, 1974 (tab.I); par Vives & al. (1975), (1982) dans la Baie Ibéro-marocaine. Dans le courant du Brésil par Berasategui & al., 2005 (p.485, tab.1) et Berasategui & al. (2006). Ces localisations atlantiques ne sont pas retenues provisoirement, de même que celles de Mazza, 1967 (p.67) en Méditerranée nord-occidentale; Soenen (1969, p.61) en Méditerranée sud-occidentale; Duran, 1963 (p.12); Seguin al., 1993 (p.23) en Mer d'Alboran; Vives (1971, p.162; 2007 p.873) en Mer Catalane; Gilabert & Moreno, 1998 (tab.1, 2) aux Baléares; Vaissière & Séguin, 1980 (p.23, tab.2) en mer Ionienne et Tyrrhénienne; Scotto di Carlo & Ianora, 1983 (p.150) dans le bassin Levantin; Scotto di Carlo & al. (1984, p.1041) en Mer Tyrrhénienne; Evans (1968, p.13) et Lapernat (1999, p.15, 55) à Malte; Pesta (1920, p.498); Regner, 1985 (p.20) et Hure & Krsinic (1998, p.20, 100) en Adriatique; V.N. Greze, 1963 a (tab.2) en Mer Ionnienne; Apostolopoulou (1972), Siokou-Frangou (1997), Moraitou-Apostolopoulou & al. (2000), Isari & al. (2006) en Mer Egée; El-Maghraby, 1965 sur la côte égyptienne; Dowidar & El-Maghraby, 1970; Salah, 1971 à Alexandrie; Scotto di Carlo & al., 1991 (p.271) en Méditerranée. Récemment Uysal & Shmeleva (2012 (p.919, Table I) rapporte cette espèce sur les côtes de Turquie (nord du bassin Levantin) avec P. sewelli.La signalisation en mer Rouge par Delalo, 1966 (1968, p.137) est possible.
Il est très difficile dans létat actuel de dénombrer le nombre de citations sûres (9) par rapport au nombre de citations (78 pour lindo-pacifique) et 270 au total. Un sérieux doute subsiste sur certaines identifications dans l'Indien.
Les dimensions données par Roe (1972): F: 4,33-6,4; M: 3,04-5,6 semblent confirmer un mélange de plusieurs espèces.
Les données fournies par le CPR (2004, p.59, fig.174) nécessitent confirmation.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 28/10/2022 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 18 juin 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
