|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Clausocalanoidea ( Superfamille ) |
|
|
|
Euchaetidae ( Famille ) |
|
|
|
Paraeuchaeta ( Genre ) |
|
|
| |
Paraeuchaeta norvegica (Boeck, 1872) (F,M) | |
| | | | | | | Syn.: | Euchäta norvegica Boeck,1872; Giesbrecht, 1892 (p.246, 264, figs.F,M);
Euchaeta norvegica : Vanhöffen, 1897 a (p.281); Giesbrecht & Schmeil, 1898 (p.40, fig.M); Mrazek, 1902 (p.515, 523, Rem.); Sars, 1902 (1903) (p.38, figs.F,M); Pearson, 1906 (p.16, Rem.); Damas & Koefoed, 1907 (p.396, tab.II, III); Farran, 1908 b (p.40); A. Scott, 1909 (p.64, 69, Rem.); Sars, 1909 b (p.16); Lysholm, 1913 (p.5); Lysholm & Nordgaard, 1921 (p.17); Sciacchitano, 1930 (p.19, figs.M); Wilson, 1932 (p.23); Nicholls, 1934 (p.31, figs.N, juv., F,M); Raymont & Gauld, 1951 (p.681, respiratory rate); Vervoort, 1957 (p.77); Lacroix, 1960 (p.11, 25); Matthews, 1967 (p.159, Table 1, Rem.); Shih & al., 1971 (p.39, 204); Matthews & Sands, 1973 (p.19, Table 4); Sargent & al., 1974 (p.217, lipid analyses); Harding, 1974 (p.141, tab.2, gut contents); Bamstedt, 1975 (p.31, weight-protein-lipid); Bamstedt & Matthews, 1975 (p.311, chemical composition); Sargent, 1976 (p.149, lipids); Bamstedt & Skjoldal, 1976 (p.63, weight-ATP-RNA); Bakke, 1977 (p.49, population dynamics); Skjoldal & Bamstedt, 1977 (p.197, adenosides); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Hopkins C., 1977 (p.723, size, development time, egg mortality); Hopkins & Machin, 1977 (p.113, spermatophore placement on genital segment); Myklebust & al., 1977 (p.283, cardiac muscle histology); Bamstedt & Holt, 1978 (p.225, feeding); Hopkins C., 1978 (p.197, male genital struture, spermatophore); Hopkins C. & al., 1978 (p.631, figs.M, structure & function of P5); Colebrook, 1978 (tab.1); Ferrari, 1978 (p.509, figs.F); Longhurst & Williams, 1979 (p.1, Table IVb, V, fig.5, 8a, vertical distribution/North Atlantic Drift); Pipe & Coombs, 1980 (p.223, figs. 1, 2, table 1, vertical distribution); Bamstedt & Skjoldal, 1980 (p.304, weight-RNA); Gagnon & Lacroix, 1982 (p.9, 12); Hopkins C., 1982 (p.91, sex ratio, seasonal variation); Buchanan & Sekerak, 1982 (p.41, Table 2: vertical distribution); Huntley & al., 1983 (p.143, Table 2, 3); Bamstedt, 1983 (p.291, RNA variation); Roe, 1984 (p.357); Tremblay & Anderson, 1984 (p.5); Sameoto, 1984 (p.213, Table 1, fig.3); 1984 a (p.767, vertical migration); Norrbin & Bamstedt, 1984 (p;47, Table 2: 3, calorific value); Skjoldal & al., 1984 (p.195, metabolic activity); Bamstedt, 1985 (p.607, excretion rate); Bamstedt & Tande, 1985 (p.259, Table 2: literature data respiration & excretion); Ward & Robins, 1987 (p.127, Table VI, reproduction); Yen, 1987 (p.283, feeding); Bamstedt, 1988 (p.15, protein content); Citarella, 1989 (p.123, abundance); Yen & Nicoll, 1990 (p.218, 222, figs.); Mauchline, 1992 a (p.2, 3); Richter, 1994 (tab.4.1a); Mauchline, 1994 a (p.561); Hays & al., 1994 (tab.1); Falkenhaug & al., 1997 (p.449, spatio-temporal pattern); Mauchline, 1998 (tab.63); 1999 (n°182, p.8, figs.F,M); Reid & Hunt, 1998 (p.310, figs.2, 3, Rem.); Nicholas & Nash R, 1999 (p.367); Halvorsen & Tande, 1999 (p.279, tab.2, 3, Rem.: p.282); Beaugrand & al., 2002 (p.1692); Beaugrand & al., 2002 (p.179, figs.5, 6); Kahle & Zauke, 2003 (p.409, metals concentration); Gislason & Astthorsson, 2004 (p.472, tab.1); G. Harding, 2004 (p.51, figs.F,M); Durbin & Casas, 2006 (p.2537, Table 2b); Hsiao & al., 2010 (p.179, Table III, trace metal concentration); Pepin & al., 2011 (p.273, Table 2, seasonal abundance); Hsiao & Fang, 2013 (p.175, Table 2: Hg bioaccumulation);
Euchaeta carinata Möbius, 1875;
Euchaete norvegica : With, 1915 (p.158, figs.F,M);
no Euchaeta norvegica : Sars, 1900 (p.58, figs.F,M);
Pareuchaeta norvegica : Sars, 1925 (p.111); Rose, 1929 (p.25); 1933 a (p.118, figs.F,M); Jespersen, 1934 (p.71, fig.18, Rem.); 1939 (p.49, Rem.); 1940 (p.28, fig.4); Lysholm & al., 1945 (p.23); Brodsky, 1950 (1967) (p.206, figs.F,M); Gundersen, 1953 (p.1, 18, seasonal abundance); Østvedt, 1955 (p.15: Table 3, p.63); Marshall & Orr, 1962 (tab.3); Fisher, 1962 (p.129, % lipids); Bary, 1963 a (p.1519, Table 1); 1964 (p.183, T- diagram-occurrences); Mazza, 1966 (p.70); Elofsson, 1966 (p.77, figs.: eye); Maclellan D.C., 1967 (p.101, 102: occurrence); Vinogradov, 1968 (1970) (p.61); Conover & Corner, 1968 (p.49, 61, respiration & nitrogen excretion); Corner & Cowey, 1968 (p.393, Table 7, respiration rate); Vidal, 1971 a (p.16, 25, figs.F,M); Beaudouin J., 1973 (p.69); Mauchline & Nemoto, 1977 (p.32, figs. F,M, juv.: pattern of integumental organs); Bamstedt, 1979 (p.135, respiratory rate, energy consumption); Kovalev & Shmeleva, 1982 (p.83); Vives, 1982 (p.291); Mikhailovsky, 1986 (p.83, Table 1, ecological modelling); Mumm, 1993 (tab.1, fig.2); Krause & al., 1995 (p.81, Fig.16, abundance, Rem.: p.131); Mauchline, 1998 (p.349: Rem., tab.20, 25, 26, 42, 45, 57, 58, 64, figs.52, 53, 74); Auel, 1999 (tab.2); Kosobokova & Hirche, 2000 (p.2029, tab.2); d'Elbée, 2001 (tabl.1); Holmes, 2001 (p.52); Auel & Hagen, 2002 (p.1013, tab.2, 3); Karnovsky & al., 2003 (p.289, Appendix 1, 2 ( auks relation/copepods); Veistheim & al., 2005 (p.382, tab.1, 2, fig.1); Blachowiak-Samolyk & al., 2006 (p.101, tab.1); Tönnesen & al., 2006 (p.21, feeding, production); Willis & al., 2006 (p.39, Table 2, advection vs changes in community structure); Conway, 2006 (p.19, copepodides 1-6, Rem.); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Raybaud & al., 2008 (p.1765, Table A1); Dalpadado & al., 2008 (p.2266, Fig.2: Md, Table 2, 3); Norrbin & al., 2009 (p.1945, Table 4, 5, abundance); Dvoretsky & Dvoretsky, 2010 (p.991, Table 2); 2011 a (p.1231, Table 2: abundance, biomass); Alvarez-Fernandez & al., 2012 (p.21, Rem.: Table 1);
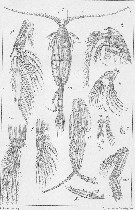
Pseudeuchaeta norvegica : Wilson, 1936 c (p.91) | | | | Ref.: | | | Wilson, 1932 a (p.65, figs.F,M); Sewell, 1948 (p.348, 496, 501); Bradford & al., 1983 (p.23); Kurbjeweit & Buchholtz, 1991 (p.168, A1 structures); Park, 1994 (p.319); 1995 (p.72, Rem.F, M, figs.F,M); Braga & al., 1999 (p.79, 84, tab.1, figs.6, 7, 8, Rem.: Biol. mol.); Boxshall & Halsey, 2004 (Rem. p.120, p.119: figs.F,M); Vives & Shmeleva, 2007 (p.666, figs.F,M, Rem.) |  issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. California, San Diego, 1995, 29. [p.179, Fig.69]. Female: a, forehead (left side); b, urosome (left); c, d, e, genital somite (left, right, ventral, respectively); f, Mx1 (first inner lobe omitted), posterior; g, P1 (anterior); h, P2 (anterior). Male: i, forehead (left); j, k, last pedigerous and genital somites (left, right, respectively); l, exopodof P1 (anterior); m, exopod of P2 (anterior); n, exopod of left 5th leg (medial); o, p, distal end of left 5th leg exopod (anterior, medial). Nota: Mx1 with 1+9+3 setae ob 1st inner lobe; 1 seta each on 2nd and 3rd inner lobes; 5 setae on basis, the outermost of which is very small; 1st endopodal segment with 7 setae; outer lobe with 9 setae, of which 2 proximal ones are shorter than the others.
|
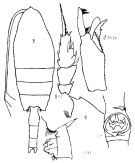 Issued from : K.A. Brodskii in Calanoida of the Far Eastern Seas and Polar Basin of the USSR. Opred. Fauna SSSR, 1950, 35 (Israel Program for Scientific Translations, Jerusalem, 1967) [p.207, Fig.110]. As Pareuchaeta norvegica. Female (from central part Arctic): habitus (dorsal); forehead (lateral); S2, exopod of P2; corner of the last thoracic segment and genital segment (lateral left side); Gf, genital segment (ventral). Male: S5Le, distal segments of left P5.
|
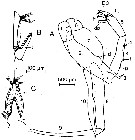 issued from : C.C.E. Hopkins, J. Mauchline & D.S. McLusky in J. mar. biol. Ass. U.K., 1978, 58. [p.633, Fig.2]. As Euchaeta norvegica. Male (from Loch Etive, Scotland): A, P5; B, 3rd exopodal segment of left P5; C, internal view of the 3rd exopodal segment of left P5. The stippled area in B and C is the concave region that receives the neck of the spermatophore. Lft P5: 1, 1st basipodal segment; 2, 2nd basipodal segment; 3, endopod; 4, 1st exopodal segment; 5, 2nd exopodal segment; E3, 3rd exopodal segment. Right P5: 6, 1st basipodal segment; 7, 2nd basipodal segment; 8, 1st exopodal segment; 9, 2nd exopodal segment; 10, endopod. d: denticle; dp: digitiform process; f, flange; l: lamella; p: peg; s: sensillum; t: tubercle. Nota: The endopod of left P5 is indistinctly divided into 2 segments and its end is flexed; the exopod consits of 3 segments, the 1st has a flange along part of its outer surface (f) with a small semi-rounded peg or denticle (p) at its proximal end and an integumental sensillum (s) at its distal end; the 2nd exopodal segment terminates in a partly toothed, internally curved, dagger-shaped lamella.; a cylindrical digitiform process (dp) is present near the distal end of this segment, it has the peculiar surface sculpturing (in below Figure 3B); the sculpturing may be indicative of a sense of touch; the 3rd exopodal segment is slender and is complex form; a tubercle with fine setae present on its internal face is located at the proximal end and groups of setae are also present at the distal end of the segment (Figure 2, B-C), the surface of the segment between the origin of the tubercle and the distal groups of setae is concave and the whole structure forms a pocket to receive an hold the neck of the spermatophore.
|
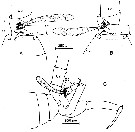 issued from : C.C.E. Hopkins, J. Mauchline & D.S. McLusky in J. mar. biol. Ass. U.K., 1978, 58. [p.634, Fig.4]. As Euchaeta norvegica. Male: The graspin of the spermatophore extruded by the ductus ejaculatorius. A-B, ventral views of the posterior region of the thorax and urosome; C, idem (lateral view); dp: digitiform process; l: lamella; E3: 3rd exopodal segment; U2: 2rd urosome segment. Nota: The spermatophore is extruded, flask end first and neck last, through the genital pore on the left ventral side of the 1st urosome segment. The sole function of the right P5 is grasping of the urosome of the female. The leftP5 transfers the spermatophore; the 3rd exopodal segment grasps the extruded spermatophore but, except for a possible role in guiding the neck of the spermatophore towards the hollow in the 3rd exopodal segment, the digitiform process and lamella do not appear to serve any obvious purpose during this operation. The digitiform process is always freely moveable and is not in contact with the adhesive material of the spermatophore attached to the exopod. The lamella, in the mating position (shown in figure 5D) lies across the posterior of the ventral face of the genital bursa of the female; it appears to act as a non-slip templet for accurate placement ofthe spermatophore on the genital openings of the female. There is still no obvious function for the digitiform process in this second operation; it may be sensory in nature, possibly sensitive to touch, and aid in the accurate grasping and placement of the spermatophore.
|
 issued from : C.C.E. Hopkins, J. Mauchline & D.S. McLusky in J. mar. biol. Ass. U.K., 1978, 58. [p.636, Fig.5, A-D]. As Euchaeta norvegica. Four possible mating positions.
|
 issued from : C.C.E. Hopkins, J. Mauchline & D.S. McLusky in J. mar. biol. Ass. U.K., 1978, 58. [p.633, Fig.3, B]. As Euchaeta norvegica. Male: B, scanning electron micrograph showing the \"thumbprint\" and well-defined ridges at the distal en of the digitiform process (dp in figures 2 & 4) on the 2nd exopodal segment of left P5. Nota: The sculpturing may be indicative of a sense of touch. A similar form of sculpturing is present in the walls of the integumental pores from which the integumental scales originate in certain genera of caridean decapods (see in Mauchline & al., 1977).
|
 issued from : Sars G.O. in An Account of the Crustacea of Norway, with short descriptions and figures of all species. Vol. IV. Copepoda Calanoida. Publ. by The Bergen Museum. 1903. [Pl. XXIV]. As Euchaeta norvegica. Female. pr = corner of the last thoracic segment; gen.S = genital segment (lateral); Urs = urosomal segment 3, anal segment and caudal rami. Nota: eggs in the ovisac dark blue. caudal setae with hairs iridescent.
|
 issued from : Sars G.O. in An Account of the Crustacea of Norway, with short descriptions and figures of all species. Vol. IV. Copepoda Calanoida. Publ. by The Bergen Museum. 1903. [Pl. XXV]. As Euchaeta norvegica. Female. M = Md; m = Mx1; mp1 = Mx2; mp2 = Mxp.
|
 issued from : Sars G.O. in An Account of the Crustacea of Norway, with short descriptions and figures of all species. Vol. IV. Copepoda Calanoida. Publ. by The Bergen Museum. 1903. [Pl. XXIV]. As Euchaeta norvegica. Male M = Md; m = Mx1; mp1 = Mx2; mp2 = Mxp..
|
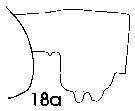 issued from : J. Mauchline in ICES Ident. Leafl. Plankton, 1999, N°182. [p.3, Fig.2: 18a]. As Euchaeta norvegica. Female (Northeast Atlantic): 18a, genital double-somite (left side).
|
 issued from : J. Mauchline in ICES Ident. Leafl. Plankton, 1999, N°182. [p.4, Fig.3: 18b]. As Euchaeta norvegica. Male (Northeast Atlantic): 18b, terminal two segments, exopodal segments of left P5.
|
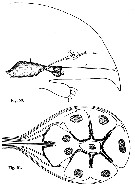 issued from : R. Elofsson in Sarsia, 1966, 25. [p.78, Figs. 50, 51]. As Pareuchaeta norvegica. Fig.50: (from Bergen, Norway) Semi-diagrammatic drawing from sections of the anterior portion of the head showing in sagittal view the spatial relationship of the organs in question; 1 = Brain; 2 = Nauplius eye; 3 = organ of Gicklhorn; 4 = X-organ. Fig.51: Semi-diagrammatic drawing from sections of the ventral cup. The arrangement of the cells is shown in dorsal view. Dotted portion of the cells indicates the rhabdomeres.
|
 issued from : R. Elofsson in Sarsia, 1966, 25. [p.81, Fig. 53]. As Pareuchaeta norvegica. Semi-diagrammatic drawing from the sections showing the axonal pathways from one lateral and the ventral cup. The sagittal view excludes part of the axones from the ventral cup to be seen.
|
 issued from : G. Harding in Key to the adullt pelagic calanoid copepods found over the continental shelf of the Canadian Atlantic coast. Bedford Inst. Oceanogr., Dartmouth, Nova Scotia, 2004. [p.51]. As Euchaeta norvegica. Female & Male. Nota: Female with appendicular setae under caudal rami almost as long as the body length.
|
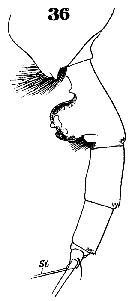 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.37, Fig.36]. As Euchäta norvegica. Female: 36, urosome (lateral).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Figs.7, 9]. As Euchäta norvegica. Female: 7, Mx1 (posterior view); 9, Mxp (distal seta from endopod).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.33]. As Euchäta norvegica. Female: 33, outer margin of Mx2.
|
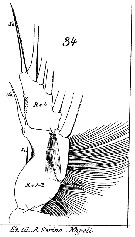 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.15, Fig.34]. As Euchäta norvegica. Female: 34, exopod of P1 (posterior view)..
|
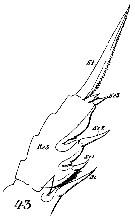 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.43]. As Euchäta norvegica. Female: 43, exopodite 3 of P2 (posterior view)..
|
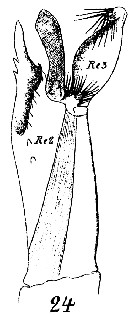 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.24]. As Euchäta norvegica. Male: 24, exopodite 2 and 3 of left P5.
|
 Issued from : C.C.E. Hopkins in J. exp. mar. Biol. Ecol., 1978, 35. [p.200, Fig.1]. Diagram of an adult male Euchaeta norvegica viewed from the dorsal aspect to show the position of the component parts of the reproductive system. Nita: The male reproductive system consists of a single testis and the left genital duct which passes posteriorly through the perivisceral cavity and opens to the exterior in the left side or the first urosome segment. The right genital duct appears to be absent. Lowe (1935) describes it as a rudimentary cord of cells lying in the right side of the pericardial floor in Calanus finmarchicus.
|
 Issued from : C.C.E. Hopkins in J. exp. mar. Biol. Ecol., 1978, 35. [p.201, Fig.2]. Diagram of the reproductive system of an adult male Euchaeta norvegica viewed from the left side. Nota: Terminology following Heberer's (1924) for the duct of heterocope: Four distinct regions can be differentiated in the structure of the wall of the genital duct, termed the vas deferens, the seminal vesicle, the spermatophore sac, and the ductus ejaculatorius.
|
 Issued from : J. Mauchline & T. Nemoto in Bull. Plankton Soc. Japan, 1977, 24 (2). [p.109, Fig.1, A]. Distributions of pores in the treated integuments of Pareuchaeta norvegica (= Paraeuchaeta norvegica) adult female from western coast of Ireland (about 55°N, 12°30'W).
|
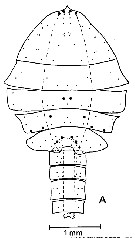 ssued from : J. Mauchline & T. Nemoto in Bull. Plankton Soc. Japan, 1977, 24 (2). [p.110, Fig.2, A]. Distributions of pores in the treated integuments of Pareuchaeta norvegica (= Paraeuchaeta norvegica) adult male from western coast of Ireland (about 55°N, 12°30'W).
|
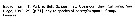 Paraeuchaeta norvegica Paraeuchaeta norvegica Female: 1 - See key to species Groups and independent species of Paraeuchaeta (p.30). 2 - Laterally, genital field facing ventrad, with large tubercular outgrowth behind genital orifice (Fig.69-c). 3 - Genital somite with transverse ridge on ventral side anterior to genital prominence (Fig.69-c).
| | | | | Ref. compl.: | | | Marshall & Orr, 1954 (p.393, Text-fig.2, egg, hatching); Conover, 1960 (p.399, Table I, respiratory rate); Wickstead, 1962 (p.546, food & feeding); Grice & Hart, 1962 (p.287, table 3); Grice, 1963 a (p.495); Itoh, 1970 a (p.8: tab.2); Bakke, 1977 (p.49, life history); Groendahl & Hernroth, 1986 (tab.1); Tiselius, 1988 (p.215, Table 1); Fransz & al., 1991 (p.9); Tiselius & Jonsson, 1997 (p.164, feeding); Weslawski & Legezynska, 1998 (p.1238); Kosobokova & al., 1998 (tab.2); CPR, 2004 (p.58, fig.171); Auel & Hagen, 2005 (p.1272, Table 2); Vestheim & al;, 2005 (p.19, vertical distribution); Manning & Bucklin, 2005 (p.233, Table 1); Hop & al., 2006 (p.182, Table 4); Valdés & al., 2007 (p.104: tab.1); Ferrari & Dahms, 2007 (p.63, 64, Rem.); Kattner & al., 2007 (p.1628, Table 1); Cabal & al., 2008 (289, Table 1); Park & Ferrari, 2009 (p.143, Table 8, Appendix 1, biogeography); Laakmann & al., 2009 (p.741, fig.2, vertical distribution); Harvey & Devine, 2009 (p.21: Table 4); Vogedes & al., 2010 (p.1471, lipid sac area vs lipid content); Bucklin & al., 2010 (p.40, Table 1, Biol mol.); Kosobokova & al., 2011 (p.29, Table 2, Rem.: Arctic Basins); Pomerleau & al., 2011 (p.1779, Table V, VI); Tang & al., 2011 (p.77, Rem.: p.61, biomass); Browman & al., 2011 (p.2653, predation); Hirche & Kosobokova, 2011 (p.2359, Table 3, abundance, biomass %); Laakmann & al., 2012 (p.535, Table 1, fig.2, Rem.: mol. Biol.); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); El Arraj & al., 2017 (p.272, table 2); | | | | NZ: | 8 + 5 douteuses | | |
|
Carte de distribution de Paraeuchaeta norvegica par zones géographiques
|
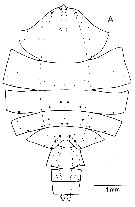
| | | | | | | | | | | |  issued from : A. Longhurst & R. Williams in J. Plankton Res., 1979, 1 (1). [p.23, Fig.8a]. issued from : A. Longhurst & R. Williams in J. Plankton Res., 1979, 1 (1). [p.23, Fig.8a].
Day/night percentage numerical profiles at India Station (59°N, 19°W) at end of March 1975.
A: As Euchaeta norvegica. B: Total copepods.
Depth in meter.
Sampling by LHPRs (Longhurst-Hardy Plankton Recorders). |
 issued from : A. Longhurst & R. Williams in J. Plankton Res., 1979, 1 (1). [p.24, Fig.8b]. issued from : A. Longhurst & R. Williams in J. Plankton Res., 1979, 1 (1). [p.24, Fig.8b].
Day/night percentage numerical profiles at India Station (59°N, 19°W) in early May 1975.
A: As Euchaeta norvegica. B: Total copepods.
Depth in meter. |
 Issued from : J.L.W. Bakke in Sarsia, 1977, 63. [p.50, Fig.1]. Issued from : J.L.W. Bakke in Sarsia, 1977, 63. [p.50, Fig.1].
Percentage abundance of stages of E. norvegica (from Korsfjorden: 60°12'N, 5°13'E) in generations.
. Females are separated into ovigerous (top) and non-ovigerous individuals.
Generations (G1-G7) are labelled at peaks of the combined stages I, II, and III.
Nota: Specimens collected every second week with the Longhurst frame sampler (0.5 mm mesh aperture), using an oblique haul of the water column.
Water circulation in this fjord is restricted by a sill at 250 m. Temperatures ranging from 5.7 to 7.9 °C within the sampling period. |
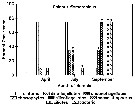 Issued from : J.L.W. Bakke in Sarsia, 1977, 63. [p.53, Fig.4]. Issued from : J.L.W. Bakke in Sarsia, 1977, 63. [p.53, Fig.4].
Average prosome lengths of E. norvegica in Korsfjorden.
Closed circles: stages I-III and femalesIV-VI. Open circles: Males IV-VI.In the equations expressing the curves fitted to the averages, y is prosome length in mm and x is stage I-VI for the females and I-V for the males.
Short horizontal lines, unbroken and broken for females and males respectively, show the values for y predicted by the equations.
The lengths of males at stage VI were not included in calculation of the regression equation since they show only a small increase following the molt from stage V, undoubtedly because the adult males do not feed. |
 Issued from : K.W. Tang, T.G. Nielsen, P. Munk, J. Mortensen, E.F. Møller, K.E. Arendt, K. Tönnesson, T. Juul-Pedersen in Mar. Ecol. Prog. Ser., 2011, 434. [p.86, Fig.8] Issued from : K.W. Tang, T.G. Nielsen, P. Munk, J. Mortensen, E.F. Møller, K.E. Arendt, K. Tönnesson, T. Juul-Pedersen in Mar. Ecol. Prog. Ser., 2011, 434. [p.86, Fig.8]
Pareuchaeta norvegica (from the continental slope off Fyllas Bank to the inner part of Godthabsfjord, SW Greenland, corresponding to stations 0 to 20) in the summer (2008).
Contour plots of biomass (mg C/m3) of all developmental stages collected from 4 to 9 strata with a multinet samples (300 µm mesh aperture)
Dots are mid-points of sampling intervals. Numbers on top are stations. Hatched area = bottom topography. |
 Issued from : K.W. Tang, T.G. Nielsen, P. Munk, J. Mortensen, E.F. Møller, K.E. Arendt, K. Tönnesson, T. Juul-Pedersen in Mar. Ecol. Prog. Ser., 2011, 434. [p.79, Fig.1] Issued from : K.W. Tang, T.G. Nielsen, P. Munk, J. Mortensen, E.F. Møller, K.E. Arendt, K. Tönnesson, T. Juul-Pedersen in Mar. Ecol. Prog. Ser., 2011, 434. [p.79, Fig.1]
Station positions along Godthabsfjord in southwestern Greenland. |
 Issued from : K.W. Tang, T.G. Nielsen, P. Munk, J. Mortensen, E.F. Møller, K.E. Arendt, K. Tönnesson, T. Juul-Pedersen in Mar. Ecol. Prog. Ser., 2011, 434. [p.81, Fig.2] Issued from : K.W. Tang, T.G. Nielsen, P. Munk, J. Mortensen, E.F. Møller, K.E. Arendt, K. Tönnesson, T. Juul-Pedersen in Mar. Ecol. Prog. Ser., 2011, 434. [p.81, Fig.2]
Contour plots of water temperature (°C), salinity, density (kg/m3) and chlorophyll a (mg/m3) along the transect of Godthabsfjord.
Distances were measured from Station o. Note the different contour line scales for different panels.
Hatched area in each panel represents bottom topography. |
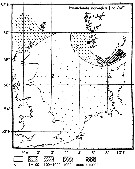 Issued from : M. Krause, J.W. Dippner & J. Beil in Prog. Oceanog., 1995, 35. [p.105, Fig.18]. Issued from : M. Krause, J.W. Dippner & J. Beil in Prog. Oceanog., 1995, 35. [p.105, Fig.18].
Horizontal distribution pattern of Pareuchaeta (= Paraeuchaeta) norvegica (individuals per m2) in the winter North Sea.
Collected by WP2-net. Numbers (ind/m3) depth-integrated, extrapoled to the bottom (but at a maximum to a depth of 500 m) and expressed as ind per m2 of water surface. |
 Issued from : S. Laakmann, M. Kochzius & H. Auel in Deep-Sea Res. I, 2009, 56. [p.745, Fig.2 a]. Issued from : S. Laakmann, M. Kochzius & H. Auel in Deep-Sea Res. I, 2009, 56. [p.745, Fig.2 a].
Vertical distribution abundance of copepodite stages C3 to C6 of Paraeuchaeta spp.
For P. barbata copepodite stages C1 to C6 are included. +: No occurrence; solid line depicts bottom profile (right axis); station 853 over the Yermak Plateau (81°22'N, 6°52'E) is separated from the other stations on the transect; other stations between East Greenland Current, the Fram Strait and the West Spitsbergen Current, into south return Atlantic Current (77°46'N-79°36'N, 6°20'E-7°29'W).
Deep-sea copepods collected from August 20 to September 16, 2006. |
 Issued from : H. Vestheim, S. Kaartvedt & B. Edvardsen in J. Plankton Res., 2005, 27 (1). [p.22, Fig.2]. Issued from : H. Vestheim, S. Kaartvedt & B. Edvardsen in J. Plankton Res., 2005, 27 (1). [p.22, Fig.2].
Vertical distribution of adult females of P. norvegica during night (filled bars) and day time (open bars) in April (a) and November (b) at 59°38' N, 10°38' S in the Oslofjord (Norway).
Error bars denote 95% confidence interval (CI) of mean number of individuals per m3.
Nota: Adult females collected closest to the surface were less pigmented than those found deeper. Pigmentation of P. norvegica was related to its depth of occurrence. Previously, P. norvegica has been described to have a variable, but always relatively high content of carotenoids, and pigmentation is especially marked at its mouthparts. Many of the individuals collected above 100 m were, however, almost comoletely transparent and lacked the characteristic of red pigmentation of individuals sampled deeper. Incubation of animals in darkness proved the degree of pigmentation to be plastic, and it changed quickly in relation to light intensity. The ability to change pigmentation can be an important adaptation for individuals selecting varying depths and light regime. A low level of pigmentation may be another adaptation to avoid visual predators (see Zaret & Kerfoot, 1975).
Non-ovigerous females were found higher in the water column than ovigerous females, and juveniles were found higher in the water column than adults. Ovigerous females expose to a greater risk by staying in the upper parts of the water column because the planktivorous fishes prefer the conspicuous ovigerous females to non-ovigerous (see Vuorinen & al., 1983; Bollens & Frost, 1991).
|
 Issued from : H. Vestheim, S. Kaartvedt & B. Edvardsen in J. Plankton Res., 2005, 27 (1). [p.23, Fig.5]. Issued from : H. Vestheim, S. Kaartvedt & B. Edvardsen in J. Plankton Res., 2005, 27 (1). [p.23, Fig.5].
Mean prosome length (a and b), dry weight (c and d), and volume of fat sac (g and h) of adult females of P. norvegica in April (left column) and November (right column).
Error bars denote 95% confidence interval (CI) of the mean. (Non-overlapping CI indicates significant different values; T-test, P = 0.005).
Nota: vertical distribution is related to body condition. In November, females collected in the upper parts of the water column differed from those collected deeper by having lower weight, slightly shorter prosome. In April, the female population is a mixture of individuals of different reproductive status, but the fay sac is smaller for shallow living individuais. The individual differences in dry wejght and the condition factor (CF: relationship between body mass and length = standard index for physiological status; see in Durbin & Durbin, 1978). For P. norvegica , there is also a relationship between dry weight and the content of lipids. It seems that the variability in vertical distribution among females, at least partly, can be explained by differences in body condition, where individuals with large energy reserves do not risk coming to the surface to feed. This hypothesis is further strenghened became the same pattern with lower values for individuals inhabiting the upper parts of the water column in April was found when measuring the fat sac. A simple linear measurement of the fat sac is shown to be a good estimation of the lipid reserves (see Miller & al., 1998).
In April, there was no difference in body condition (CF) in relation to depth of occurrence for non-ovigerous females. A non-ovigerous female could be both a recently spent female and a female just about to start reproducting. Since a lot of energy is transferred from the female to her eggs during maturation (see Bamstedt, 1975; Blades-Eckelbarger, 1991), it is not unexpected to find a great amount of individual variation in body condition (CF) within this group. |
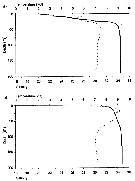 Issued from : H. Vestheim, S. Kaartvedt & B. Edvardsen in J. Plankton Res., 2005, 27 (1). [p.20, Fig.1]. Issued from : H. Vestheim, S. Kaartvedt & B. Edvardsen in J. Plankton Res., 2005, 27 (1). [p.20, Fig.1].
Temperature (stippled line) and salinity ( solid line) profiles in April (a) and November (b) at 59°38' N, 10°38' S in the Oslofjord (Norway).
Nota: In April, the upper 50 m was relatively cold. Temperature dropped from 6.2 °C in the surface to 5.3 °C at 14 m depth. Further down, temperature increased until it was stable at ± 7.3°C from 30 m to the bottom (200 m). Salinity increased from 20.4 in the surface till 34.0 at 40 m, and from 80 mm it was stable at 34.4 all the way down.
In November, surface temperature was 6.6 °C and there was a temperature maximum of 9.2 °C at 18 m. Temperature was stable at ± 7.1 °C from 90 m to the bottom. Salinity increases from 31.5 in the surface to 34.6 at 80 m. Further down, salinity did not change markedly. |
| | | | Loc: | | | Arct. (Baffin Sea, Nansen Basin, Laptev Sea, Amundsen Basin, Makarov Basin), Norway Sea, Wyville Thomson Ridge, Spitzbergen, Barents Sea, Franz Josef Land, W Baffin Bay, W Greenland, Godthabsfjord, Davis Strait, Greenland Sea, Fram Strait, Kongsfjorden, Kosterfjorden, Iceland, Faroe Is., Norvegian coast, Malangen fjord, Bergen, Korsfjorden, Raunefjorden (all the year), Oslofjord, Loch Etive (Scotland), North Sea, Skagerrak, off Rockall Is., Newfoundland, off SW Ireland, W Ireland, Ireland Sea, Fairlie Channel, Loch Fyne, off Bermuda: Station "S" (32°10'N, 64°30'W), off SE & E Cape Cod, Georges Bank, Chesapeake Bay, G. of Maine, Woods Hole, off SE Nova Scotia, Nova Scotia, Northumberland Strait, Baie des Chaleurs, G. of St. Lawrence, upper St. Lawrence estuary, Rimouski, off Morocco-Mauritania, off Madeira, off Portugal, off W Cape Finisterre, S Bay of Biscay, Azores, Medit. (W Basin, Ligurian Sea), South Africa (E) (in De Decker & Mombeck, 1964, p.13), [ Philippines, Japan (off Honshu), off Hokkaido, off Queen Charlotte Is., Aleutian Is. ( Aika Is.), SW Bering Is. (références Pacific only in C. B. Wilson, 1950, p.283) ], W Costa Rica (in Morales-Ramirez & Suarez-Morales, 2008 (p.520) | | | | N: | 164 | | | | Lg.: | | | (3) F: 8,5-7,7; M: 6,8-6; (7) F: 8,86; M: 6,34; (22) F: 7-11; M: 5,5-7; (45) F: 8,5-7,5; M: 7,5-6,5; (47) F: 7; M: 5,45; (65) F: ± 8; M: 7; (377) F: 8,79; (1303) F: 7,45; M: 6,14; {F: 7,00-11; M: 5,45-7,50}
The mean female size is 8.209 mm (n = 11; SD = 1.1413), and the mean male size is 6.423 mm (n = 10; SD = 0.6699. The size ratio (male : female) is 0.786 (n = 3; SD: 0.0906) or ± 79 %. In samples, the sex ratio (female : male) is temporary 1.14.
(1317) Prosome length female): 4,78-5,72 (with egg mass); 5,12-5-79 (ripening); 5,34-5,70 (non-ovigerous) | | | | Rem.: | épi à hadopélagique. 230-580 m (Pipe & Coombs, 1980, Arctic (60°N).
Sars (1903) ne reconnaît pas la synonymie entre E. glacialis Hansen, 1887 et cette espèce.
Park (1995) ne retient pas les localisations dans le Pacifique. De même sa présence à l'est de l'Afrique du Sud est à confirmer.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 24/10/2022 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 17 juin 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
