|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Arietelloidea ( Superfamille ) |
|
|
|
Heterorhabdidae ( Famille ) |
|
|
|
Heterorhabdus ( Genre ) |
|
|
| |
Heterorhabdus tanneri (Giesbrecht, 1895) (F,M) | |
| | | | | | | Syn.: | Heterochaeta tanneri Giesbrecht, 1895 c (p.259, pl.4, figs.5-6:M);
Heterorhabdus proximus Davis, 1949 (p.57, figs.F,M); Shih & al., 1971 (p.148);
no Bradford, 1971 a (p.126, figs.F,M);
Heterorhabdus pilosus Tanaka, 1953 (p.134) | | | | Ref.: | | | Giesbrecht & Schmeil, 1898 (p.115, Rem.M); Brodsky, 1950 (1967) (p.356, figs.F,M); Tanaka, 1964 a (p.11, figs.F,M); Arashkevich, 1969 (p.705, figs.F); Morris, 1970 (p.2301, 2316, Rem.); Vaupel-Klein, 1970 (p.38, fig.F, Rem.); Minoda, 1971 (p.40); Heptner, 1971 (p.151, Rem.); Bradford, 1971 a (p.124, Rem.); Kos, 1976 (Vol. II, figs.F, M, Rem.); Gardner & Szabo, 1982 (p.374, figs.F, M); Chihara & Murano, 1997 (p.819, Pl.118,121: F,M); Bradford-Grieve & al., 1999 (p.883, 944, figs.F,M); Park, 2000 (p.138, figs.F,M, Rem.); Sedova & Grigoryev, 2013 (p.11, figs.F,M, Rem.: morphologic variation); Takenaka & al., 2013 (p.201, DNA sequences vs luciferase); Lewis A., 2014 (p.1200, Table I, II, figs. labrum-paragnaths) | 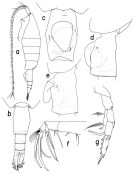 issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. California, San Diego, 2000, 31. [p.264, Fig.112]. Female: a, habitus (left side); b, urosome (dorsal); c, d, genital somite (ventral, left, respectively); e, genital somite with operculum pratially open (left); f, left Mx2 (posterior); g, P5 (posterior). Nota: Mx2: Posterior subterminal spine of 4th lobe 58% as long as 2nd saberlike spine which is a little longer than 1st saberlike spine; falcate spine of 5th lobe about 1.3 times length of saberlike spine, with proximal 52% of medial margin serrated with spinules; falcate spine of 6th lobe with proximal 68% of medial margin serrated with spinules.
|
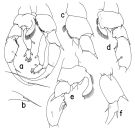 issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. California, San Diego, 2000, 31. [p.265, Fig.113]. Male: a, P5 (anterior); b, distal end of exopod of right P5 (lateral); c, basipod of left P5 (anterior); d, e, right P5 (with endopod omitted), posterior, anterior, respectively; f, exopod of left P5 (anterior).
|
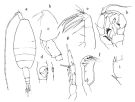 issued from : O. Tanaka in Publs Seto Mar. Biol. Lab., 1964, XII (1). [p.12, Fig.178]. Female: a, habitus (dorsal); b, forehead (lateral left side); c, last thoracic segment and genital somite (lateral left side); d, genital somite (ventral) and margin left side); e, Mx2; f, Mxp; g, P5. Nota: The urosome segments and furca are in the proportional lengths as 49:11:10:7:23 = 100. A1 extends to the end of the furca (it over-reaches the furca by 2 or 3 segments when fully reflexed). Male: h, P5; i, 1st and 2nd segment of exopod of left P5. Nota: The urosome segments and furca are in the proportional lengths as 22:20:19:12:7:20 = 100. The mouth parts and the 1st to 4th swimming legs as in the female.
|
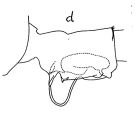 Issued from : J.C. von Vaupel-Klein in Zool. Verh. Leiden, 1970, 110. [p.32, Fig.15,d]. Female: d, genital complex (lateral left side). Nota: The caudal borders of the urosomal somites were found to have toothed edges dorsally and dorso-laterally; this detail is not mentioned by Brodsky (1950).
|
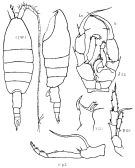 Issued from : K.A. Brodskii in Calanoida of the Far Eastern Seas and Polar Basin of the USSR. Opred. Fauna SSSR, 1950, 35 (Israel Program for Scientific Translations, Jerusalem, 1967) [p.356, Fig.251]. Female (from NW Pacif.): habitus (dorsal and lateral left side); MRi, Md (biting edge, right side view); Mp2, Mxp; S5, P5. Male: S5, P5 (Ri = right leg; Le = left leg).
|
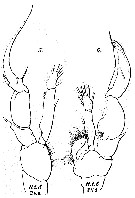 issued from : W. Giesbrecht in Bull. Mus. comp. Zool. Harv., 25 (12). [Taf. IV, Figs.5-6]. As Heterodchaeta tanneri. Male (from 35°N, 125°W): 5, left P5; 6, right P5.
|
 issued from : C.C. Davis in Univ. Wash. Publs Biol., 1949, n. ser. 14. [Pl.10, Figs.119-125]. As Heterorhabdus proximus. Female (from NE Pacific): 124, P5. Nota : Head and 1st thoracic segment nearly , but not completely, separated (there is a marked cervical groove). Last segment of prosome with rounded corners. Metasome about three and one-half times the length of the urosome. Genital segment, which bears a large ventral swelling, is as long as the following 3 segments together, plus about ½ the length of the caudal rami ; 2 nd urosomal segment a little more than 1/3 the length of the genital segment, and the same length as the two following segments together ; the 3 rd segment 3 times the length of the anal segment ; caudal rami asymmetrical, the left one longer than the right one. A1 25-segmented, reach slightly beyond the end of the caudal rami. Mxp with the seta in the middle of the inner border of the 1 st basal segment, heavy and long (it is longer than in H. papilliger. Legs with a structure similar to those of H. abyssalis. P5 with a structure between that of H. papilliger and H. abyssalis ; the setae on the inner border of the 1 st and 2 nd endopodal segments are shorter and thinner than the setae of the 3 rd segment (but they are not as short and thin as in case of abyssalis. Male: 119, right P5; 120, left P5; 122, Mx2; 123, Mxp; 125, P3. Nota : Articulation between the head and the 1st thoracic segment only very faintly indicated. Urosome nearly 1/3 the length of the metasome. From a lateral view, measured along the dorsal edge, the 2 nd urosomal segment is the longest ; the 1 st and 3 rd segments are equal in length and 5/6 of the length of the 2 nd ; the 4 th segment is only 5/8 the length of the 2 nd ; anal segment very short and only about 1/5 the length of the 2 nd. Left caudal ramus somewhat longer than the right, and the 2 nd from the inner seta that it bears is heavier than the others, anda t least one and one-half times the length of the entire animal. A1 reach about the middle of the urosome ; right A1 25-segmented, left one geniculate, 21-segmented. P5 rather similar to those of H. abyssalis ; the spine at the end of the 3 rd exopodal segment of the right le gis shorter than in abyssalis, and the shape of the protrusion on the inner border of the 2 nd exopodal segment is different.
|
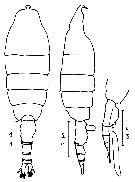 Issued from : N.A. Sedova & A.S. Grigoryev in Zool. Zh., 2013, 92 (1). [p.13, Fig.1]. Female (from Okhotsk Sea, NW Pacific): 1-2, habitus (dorsal and lateral, respectively); 3, urosome, with spermatophore (lateral). Scale bars = 1 mm.
|
 Issued from : N.A. Sedova & A.S. Grigoryev in Zool. Zh., 2013, 92 (1). [p.14, Fig.2]. Female: 1, A1; 2, proximal segments of right A1. Scale bars: 1 mm (1); 0.5 5 mm (2) Nota: A1 with 25 free segments
|
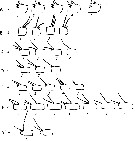 Issued from : N.A. Sedova & A.S. Grigoryev in Zool. Zh., 2013, 92 (1). [p.15, Fig.3]. Variations of armature on antennular segments. A, segment 1; B, segment 8. C-G, 9th to 12th segments from left A1 male, respectively; G, 17th segment from left A1 male.
|
 Issued from : N.A. Sedova & A.S. Grigoryev in Zool. Zh., 2013, 92 (1). [p.16, Fig.4]. Female & Male: 1, A2; 2, Mx1: 3, Md (palp); 4, Md (gnathobase); 5, Mx2; 6, Mxp. Scale bars: 0.1 mm (1, 2); 0.2 mm (3-6).
|
 Issued from : N.A. Sedova & A.S. Grigoryev in Zool. Zh., 2013, 92 (1). [p.17, Fig.5]. 1, Variations of setae of proximal setae on Mx1; 2, exopod of A2; 3, Seta on Mx2; 4, Setae on endopod of Mx2: 5, abnormal proximal sesegment on Mxp; 6, basipod segment 1 of Mxp; 7, mrophological variation on Mx1.
|
 Issued from : N.A. Sedova & A.S. Grigoryev in Zool. Zh., 2013, 92 (1). [p.19, Fig.6]. Female & Male: 1-4, P1 to P4, respectively; 5, basipod of P4; 6, abnormal basipodal segment 1 of P3. Scale bar = 0.5 mm.
|
 Issued from : N.A. Sedova & A.S. Grigoryev in Zool. Zh., 2013, 92 (1). [p.20, Fig.7]. Variations of basis of swimming legs: 1, basis of P1; 2, basis of P5 male; 3, basis of left P5 male; 4, processes on basis of P2-P4.
|
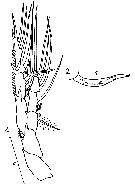 Issued from : N.A. Sedova & A.S. Grigoryev in Zool. Zh., 2013, 92 (1). [p.21, Fig.8]. Female: 1, P5; 2, spine on the inner margin of exopod segment 2. Scale bar: 0.2 mm.
|
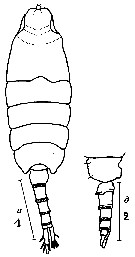 Issued from : N.A. Sedova & A.S. Grigoryev in Zool. Zh., 2013, 92 (1). [p.13, Fig.1]. Male: 1, habitus (dorsal); 2, last thoracic segment and urosome (lateral). Scale bars: 1 mm.
|
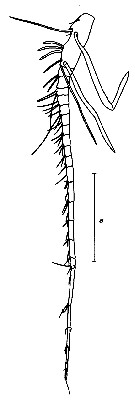 Issued from : N.A. Sedova & A.S. Grigoryev in Zool. Zh., 2013, 92 (1). [p.14, Fig.2]. Male: A1. Scale bar = 1 mm.
|
 Issued from : N.A. Sedova & A.S. Grigoryev in Zool. Zh., 2013, 92 (1). [p.21, Fig.8]. Male: A, A1; B, forms of proximal process on the exopodite 2 of right P5; C-D, variations of segments of the right and left P5. Scale bars: 0.2 mm. mm.
|
 Heterorhabdus tanneri Heterorhabdus tanneri female: 1 - See key to species groups of Heterorhabdus: ''abyssalis'' Group (p.90, 114). 2 - Genital somite without a conical projection mediodorsally (Fig.112a-a, d). 3 - Laterally, genital somite without a tubercular outgrowth posteriorly on dorsal margin (Fig.112-e, d). 4 - Laterally, genital somite with a tubercular outgrowth posteriorly on ventral margin (Fig.112-e). 5 - Laterally, genital operculum short of reaching posterior end of somite (Fig.112-d).
|
 Heterorhabdus tanneri Heterorhabdus tanneri male: 1 - See key to species groups of Heterorhabdus: ''abyssalis'' Group (p.90, 114). 2 - Basis of left P5 without a well-developed inner lobe (Fig.113-c). 3 - Basal inner lobe of right P5 armed with normal bristles (Fig.113-d). 4 - 3rd exopodal segment of right P5 with a short terminal lobe, equal or less than 1/3 length of terminal spine (Fig.113-e). 5 - Basal lobe of right P5 arising from anterior side of segment (Fig.113-a).
|
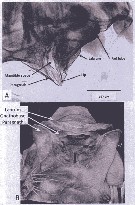 Issued from : A. Lewis in Crustaceana, 2014, 87 (10). [p.1218, Fig.11]. Heterorhabdus tanneri (Claus, 1863): A, lateral view of labrum-paragnath complex including anterior labral lobe (Ant lobe) but without mandible; B, confocal: ventral view showing paragnaths, claw on each gnathobase and labrum. See: http://booksandjournals.brillonline.com/content/journals/15685403. Nota: Relative to the body axis, the average angle of the labral-paragnath complex is + 30°; the complex faces down and well forward. The anterior margin of the labrum proper is located a very short distance behind the first antenna, about 7% of the total body length. In lateral view, the labrum and paragnath are similar length and form a beak-like structure; in ventral view however, both are flatter and do not appear beak-like. In ventral view, the distal margin of the labrum bears some very small spinules and a small, roughened surface. The anterior labral lobe is short, forming a continuous extension of the labrum. The length of the complex is slightly over 11% of the total body length, 15% with the anterior labral lobe. The length of the labrum base is 40% of the complex length. The height of the labrum is slightly less than twice the length (without anterior labral lobe). The length of the paragnath base is 60% of the complex length. The height of each paragnath is slightly more than 1.2 times the length.
|
 Issued from : A. Lewis in Crustaceana, 2014, 87 (10). [p.1219, Fig.12, B]. Heterorhabdus tanneri (Claus, 1863): B, gnathobase. See: http://booksandjournals.brillonline.com/content/journals/15685403. Nota: The unique gnathobase armature found in H. tanneri (comparatively to Eucalanus bungii, Metridia lucens, Neocalanus cristatus, Paraeuchaeta elongata), two spike-like teeth and a claw-likeventral tooth, along with the reported labral secretory cells found in members of the genus (Nishida & Ohtsuka, 1996) make it a very unique carnivore although the gnathobase and paragnaths could also be useful with larger particulate detritus. H. tanneri belongs to a group that is suggested to use venom-like injections to disable their prey (see Nishida & Ohtsuka, 1996).
|
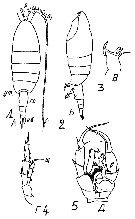 Issued from : M.S. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. II, 1976. After Brodsky, 1950. Female: 1-2, habitus (dorsal and lateral, respectively); 3, Md (side not specified; 4, P5. Male: 5, P5.
| | | | | Ref. compl.: | | | Cleve, 1904 a (p.191); Sewell, 1948 (p.519, 552, 558, 573); Minoda, 1958 (p.253, Table 2, abundance); Furuhashi, 1966 a (p.295, vertical distribution vs mixing Oyashio/Kuroshio region, Table 8); Fleminger, 1967 a (tabl.1); Vinogradov, 1968 (1970) (p.257); Shih & al., 1971 (p.148); Rudyakov, 1986 (tab.2); Mackas & Anderson, 1986 (p.115, Table 2, as tyanneri); Hattori, 1991 (tab.1, Appendix); Shih & Marhue, 1991 (tab.2, 3); Suarez-Morales & Gasca, 1998 a (p.110); Yamaguchi & Ikeda, 2000 b (p.29, vertical distribution); Yamaguchi & al., 2002 (p.1007, tab.1); Park, W & al., 2004 (p.464, tab.1); Sedova & Grigoriev, 2005 (p.112); Lavaniegos & Jiménez-Pérez, 2006 (p.145, tab.2, 3, Rem.); C.-Y. Lee & al., 2009 (p.151, Tab.2); Galbraith, 2009 (pers. comm.); Park & Ferrari, 2009 (p.143, Table 8, biogeography); Homma & Yamaguchi, 2010 (p.965, Table 2); Homma & al., 2011 (p.29, Table 2, 3, abundance); Abe & al., 2012 (p.100, temporal & vertical changes, gut content); Takenaka & al., 2012 (p.1669, fig.3, Table 1, bioluminescence); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Ohashi & al., 2013 (p.44, Table 1, Rem.); Coyle & al., 2014 (p.97, table 3); Chiba S. & al., 2015 (p.968, Table 1: length vs climate) | | | | NZ: | 5 + 1 douteuse | | |
|
Carte de distribution de Heterorhabdus tanneri par zones géographiques
|
| | | | | | | | | | Loc: | | | NW Taiwan (rare), Japan (Sagami Bay, off Sanriku, off E Hokkaido), Station H, Station Knot, Okhotsk Sea, Kuril-Kamchatka, S Bering Sea, S Aleutian Basin, S Aleutian Is., G. of Alaska, Icy Strait, Station "P", NE Pacif. (Queen Charlotte Is., off Cape Flattery), Pacif. (central subarctic), Portland Inlet, British Columbia, Fjord system (Alice Arm & Hastings Arm), off California, W Baja California, [Cuba (in Campos, 1982), ? [South Africa (E) (in Cleve, 1904 a)], Antarct. (Weddell Sea)]
Type locality: 35°N, 125°W. | | | | N: | 41 | | | | Lg.: | | | (22) F: 4,1-4; M: 3,9-3,6; (23) F: 4,65-4,2; M: 4,25-3,35; (47) M: 3,6; (59) F: 3,7-3,2; M: 3,4-3,2; (92) F: 4,1-3,47; M: 3,78-3,4; (121) F: 4,01; M: 3,85-3,75; (205) F: 4,9-4; M: 3,85; (208) F: 4,4-3,9; M: 4-3; (824) F: 4,41-3,08; M: 4,12-3,04; (1232) F: 4-4,65; M: 3,35-4,25; {F: 3,08-4,90; M: 3,00-4,25}
Chiba S. & al., 2015 (p.971, Table 1: Total length female (June-July) = 3.8 mm [optimal SST (°C) = 5.6].
| | | | Rem.: | "Abyssalis" Group.
méso-bathypélagique.
Après consultation des types, Morris (1970, p.2316) estime que cette espèce est synonyme de Heterorhabdus proximus Davis, 1949.
Voronina & Kolosova (1999, p.71) signalent cette espèce en Mer de Weddell, ce qui paraît douteux.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 19/07/2021 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2025. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 16 août 2025] © copyright 2005-2025 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
