|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
| |
| | | |
| Acartiidae Sars, 1903 ( Diaptomoidea ) | | Ref.: | Sars, 1903 (p.147); Gurney, 1931 (p.85, 214); Rose, 1933 a (p.267); Brodsky, 1950 (1967) (p.83, 418); Gonzalez & Bowman, 1965 (p.255) ; Andronov, 1974 a (p.1005); Bradford, 1976 (p.162, Rev.); Björnberg & al.,1981 (p.661); Bowman & Abele, 1982 (p.9); Razouls, 1982 (p.596); Brodsky & al., 1983 (p.142,145); Zheng Zhong & al., 1984 (1989) (p.257); Sazhina, 1985 (p.119, N); Mauchline, 1988 (p.715, 740); Huys & Boxshall, 1991 (p.460); Yoo & al., 1991 (p.257, F,M Keys); Razouls, 1993 (p.307); Madhupratap & al., 1996 (p.863, Table 5: %/copepods); Chihara & Murano, 1997 (p.669, G. & spp. Keys); Bradford-Grieve, 1999 (n°181, p.2); Barthélémy, 1999 a (p.56, 58, Rem.); Bradford-Grieve & al., 1999 (885, 901, 904, 961: Genera Key); Bradford-Grieve, 1999 b (p.213, Def., Rem.); Ohtsuka & Huys, 2001 (p.461); Boxshall & Halsey, 2004 (p.12; 49; 50: Def.; p.52: Genera Key); Mulyadi, 2004 (p.136, Def.); Vives & Shmeleva, 2007 (p.401, Genera Key); Hirai & al., 2020 (p.1, Fig.4: metabarcoding)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm  | | Rem.: | Type-genus: Acartia Dana, 1846. Total: 5 G. : Acartia, Acartiella, Paracartia, Paralabidocera, Pteriacartia.
The genus Paracartia is not always accepted by all the authors.
For Madhupratap & Haridas (1994, p.74), the subgenera created by Steuer (1923) has to be reevaluated. Steuer mainly used the presence/absence and shape of spines on pediger 5 and A1 to distinguish the subgenera In the litterature and figures on mouthparts of the subgenera show that their structures are similar and variations occur only in certain details of A1 and cutting edge of the Md. Further the presence/absence/shape of spines on pediger 5 are variable within some of the subgenera (e.g. Paracartia, Acanthacartia. The division of Acartia into arostratae and rostratae itself may not be tenable (Bowman, 1985). Hence the status and validity of the subgenus Euacartia and the other subgenera is uncertain in the family Acartiidae.
For Barthélémy (1999 has, p. 59) the genital structures do not justify the creation of 6 sub-genera. The species of the various sub-genera are distributed in a variable way in 3 of 4 subgroups of species corresponding to four variants of the model acartiidae (cf. Fig.5, Plan 9). Account was already held by the absence of significant morphological criteria of distinction of the sub-genera, the characteristics of the genital area do not justify more to keep the sub-genera.
Definition from Bradford-Grieve (1999 b, p.213) :
Female :
- A single eye present dorsally.
Cephalosome and pediger segment 1 separate, pedigers 4 and 5 always fused.
- Posterolateral corners of the prosome rounded or pointed.
- Urosome 3-segmented.
- anal operculum may be on anal segment or anus may open between the last two urosome segments into a dorsal grove on anal segment.
- Caudal rami generally slightly asymmetrical, short or long, sometimes fused with anal segment, with 6 setae.
- Without a rostrum with or without paired filaments.
- Upper lip large, prominent and trilobed ;
- A1 17-22-segmented, many segments with long setae.
- A2 endopod 3-segmented ; basis (with 1 seta) and endopod segment 1 fused, the resulting segment bears 9 setae arranged in a proximal group of 8 and 1isolated distal seta, the 8 proximal setae are interconnected at their bases by tiny tendinous strands extending from a single muscle inserted adjacent to last seta, enabling 8 setae to be moved as a single unit (Acartia) ; endopod segment 2 with 6-9 distal setae ; small terminal segment with 7 terminal setae ; exopod with reduced number of segments : 4-segmented with 1, 2, 2, 3 setae respectively.
- Md with well developed blade with a large separate tooth on one border ; palp basis with 1-2 setae ; endopod 1-segmented with 2+8-9 setae ; exopod 5-segmented with 1, 1, 1, 1, 2 setae respectively.
- Mx1 with reduced lobes and setation ; inner lobe 1 (ptaecoxal arthrite) short with about 6-8 spines and setae, inner lobes 2 and 3 and basis with 0, 3, 1 setae ; endopod apparently absent ; exopod with 2+5 setae ; outer lobes 1 and 2 with 8-9 and 1-0 setae.
- Mx2 lobes 1-5 with 3-4, 2-3, 2-3, 1-3, 1-2 setae respectively ; endopod with 4-5 setae.
- Mxp reduced and highly modified, coxa with 5-6 long setae ; basis with 1 short thick seta ; endopod with fused segments with 4-5 short thick setae.
- Swimming legs P1-P4 slender and delicate with long natatory setae ;
- Distolateral borders of exopod segments of P2-P4 expanded into a conspicuous tooth ; articulated spines are not present in this position.
- P1 basis may have a small outer edge spine or seta.
- Terminal exopod spine of P2-P4 with outer edge teeth.
- Female P5 usually uniramous, small, symmetrical, 3-segmented ; coxa and coupler may be fused or coxa and basis and the coupler (intercoxal sclerite) may be fused (Paracartia) ; basis with an outer seta ; exopod in the form of a spine or elongate seta ; small endopd present in Acartiella.
Male :
- Urosome 5-segmented.
- A1 usually prehensile on the right side only, with 12-18 ? segments ; middle segments may be greatly inflated.
- Mouthparts more or less the same in both sexes.
- P5 uniramous, asymmetrical, attached to a plate formed from fusion of coxa and coupler ; basis usually asymmetrically developed with an outer distal seta ; left leg with 2-segmented exopod (3-segmented in Paracartia africana), exopod segment 2 with variously decorated tip ; right leg usually a 3-segmented exopod, exopod segments 1 and 2 often with inner processes ; exopod segment 3 opposed to the process on exopod segment 2 to form a type of subchela.
Diagnosis from Boxshall & Halsey (2004, p.50) :
- Cephalosome and pedigerous somite 1 separate, pedigerous somites 4 and 5 fused.
- Posterolateral angles of prosome rounded or pointed.
- Urosome 3-segmented in female with 2 free abdominal somites
- Genital apparatus of female with paired gonopores and copulatory pores situated in paired slits (not concealed beneath common operculum), typically located either adjacent to midline on ventral surgace of genital double-somite, or located ventrolaterally on lateral margins of double-somite, as in Paralabidocera ; paired seminal receptacles usually present, rarely absent, as in Acartiella.
- Urosome 5-segmented in male ; single genital aperture located ventrolaterally at posterior rim of genital somite on right side.
-Caudal rami ofthen slightly asymmetrical, with up to 6 setae ; sometimes fused to anal somite.
- Rostrum represented by paired rostral filaments or absent.
- Nauplius eye present.
- A1 17 to 22-segmented in female with several compound segments in proximal part ; distally segments XXIV, XXV, XXVI all typically free, not fused to apical double-segment (XXVII-XXVIII).
- A1 geniculate on right side only in males ; secondary segmental fusions distal to geniculation typically XXI-XXIII, XXIV-XXVI and XXVII-XXVIII, or XXIV-XXVIII.
- A2 biramous, coxa with 1 seta ; basis and 1st endopodal segment forming elongate allobasis bearing 6 to 8 proximal and 1 distal setae along inner margin ; 2 nd endopodal segment elongate bearing 6 to 9 setae ; 3rd endopodal segment short bearing 5 setae plus 2 fused setae (representing vestigial 4th segment ; exopod short 4 or 5-segmented, setation formula 1, 2, 2, 3 or 2, 1, 1, 1, 3. Endopod apparently reduced to short segment bearing 9 setae in Acartiella.
- Labrum large, trilobate.
- Md comprising coxa with well developed gnathobase and disyal palp consisting of basis with 1 seta ; indistinctly 2-segmented endopod and 5-segmented exopod ; endopodal segments 1 and 2 with 2 and 8 or 9 setae respectively ; exopodal setation formula 1, 1, 1, 1, 2. Palp often with setation reduced, or entire palp sometimes vestigial or absent.
- Mx1 with weakly developed praecoxal arthrite bearing about 6 to 8 elements ; coxa with endite bearing 3 setae and with 8 or 9 setae on epipodite ; basis, endopod and exopod fused, armed with 1 inner and 1 outer basal seta, 5 distal detae representing endopod, and 2 outer setae representing exopod ; setation sometimes further reduced.
- Mx2 with praecoxa and coxa separate, setation formula of endites variable ¾, 2/3, 2/3, 1/2/3 ; basis with 2 setae ; endopod 4-segmented, segmental setation formula 1, 1, 1, 2.
- Mxp 4-segmented ; robust syncoxa and reduced distal part of limb ; syncoxa with up to 5 well developed spinulate setae ; basis and 2-segmented endopod bearing 1, 3, 2 short setae ; proximal endopodal segment with indication of subdivision into 3 segments ; often with setation reduced.
- Swimming legs P1 to P4 biramous, typically with 3-segmented exopods zand 2-segmented endopods due to fusion of 1st and 2 nd endopodal segments in P2 to P4/
- Outer distal angles of exopodal segments of P2 to P4 with large spinous processes.
- Inner seta on basis of P1 absent.
Key to genera after Boxshall & Halsey (2004, p.52) :
1 Female P5 biramous ; female and male left A1 21 or 22-segmented
.. 2.
1 Female P5 uniramous ; female and male left A1 18 or 19-segmented
. 3.
2 Endopod of A2 reduced, about as long as exopod ; female A1 21-segmented
Acartiella.
2 Endopod of A2 elongate, much longer than exopod ; female A1 22-segmented
.. Paralabidocera.
3 Posterolateral angles of prosome expanded into wing-like processes ; plumose seta of P5 shortyer than terminal claw
. Paracartia.
3 Posterolateral angles of prosome rounnded, not expanded ; female P5 reduced to minute lobe bearing single apical seta
Pteriacartia.
3 Posterolateral angles of prosome rounded or pointed, not wing-like ; plumose seta of P5 typically longer than terminal claw
. Acartia. After Boxshall & Halsey (2004, p.52) acartiids live near-seurface waters in estuarine and coastal habitats, especially embayments; only two species Acartia danae and Acartia negligens primarily inhabit open oceanic waters. Some species may occir in continental waters that are connected at least temporarily to marine waters (Dussart & Defaye, 2002). | 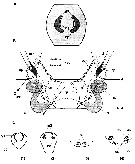 issued from : R.-M. Barthélémy in These Doc. Univ. Provence (Aix-Marseille I), 1999. [Fig.5]: Female: Schéma of the genital area in modele Acartiidae. A: external ventral view. gn = gonopore.gs = genital slit; cp = copulatory pore; asterisk = cuticular shutter. B: internal dorsal view. ap = apodemem; m2 = opercular muscle ; ed = egg-laying duct; gn = gonopore or gonoporal slit; gs = genital slit; sd = seminal ductsr = seminal receptacle; cp = copulatory pore. C: Sectional drawings of genital structures. 1, 2, frontal cross-frontal; 3-4, cross-section |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod diversity. The Ray Society, 2004, No 166, Part 1. [p.52]. Genera Key. |
 Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.214]. Spine and seta formula for swimming legs P1 to P4. Spine = Roman numeral; Arabic numeral = seta. The element on the outer margin of any segment is given first, separated by a hyphen from the inner margin element. Nota: Basis of P1 may have a small outer edge spine or seta; terminal exopod spine of legs P2-P4 with outer edge teeth. Female P5 usually uniramous, small, symmetrical, 3-segmented, basipod 1 (= coxa) and coupler may be fused or basipods 1 and 2 and the coupler may be fused ( Paracartia/em>); basipod 2 (= basis) with an outer seta; exopod in the form of a spine or elongate seta; small endopod present in Acartiella. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, Part I, No 166. [p.50]. Armature of swimming legs P1 to P4. Nota: Swimming legs P1 to P4 biramous, typically with 3-segmented exopods and 2-segmented endopods due to fusion of 1st and 2nd endopodal segments in P2 to P4. Outer distal angles of exopodal segments of P2 to P4 with large spinous processes. Inner seta on basis of P1 absent; spine and setal formula based on Acartia longiremis (Lilljeborg) but indicating range within family. - Setation sometimes reduced; 3 inner setae present on endopodal segment 1 of P4 in Paralabidocera. - P5 female comprising common transverse plate, forming partial fusion of coxae and intercoxal sclerite, and basis armed with outer seta derived from basis; exopod either a well defined segment bearing spinous processes (as in Paralabidocera) or reduced to a curved spine (as in Acartia). Usually uniramous, lacking endopod, btut endopod present as small, tapering segment in Paralabidocera and Acartiella. - Male P5 asymmetrical; intercoxal sclerite partly sefined; right leg uniramous, comprising more or less fused coxa and basis, armed with outer seta, plus 1 to 3-segmented exopod; left leg uniramous, comprising more or less fused coxa and basis, armed with outer seta, plus 1 or 2-segmented exopod. - Eggs released into water. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, Part I, No 166. [p.51, Fig.2]. Acartiidae. A, Acartia clausi habitus female (dorsal view); B, female P5; C, male P5; D, Mxp; E, A2. [Sars, 1903a: A-C; Huys & Boxshall, 1991: D-E]. | | | | | | Syn.: | Dias Lilljeborg, 1853; Claus, 1863 (p.191); Brady, 1878 (p.50) | | Ref.: | Brady, 1883 (p.72); Giesbrecht, 1892 (p.75, 506); Giesbrecht & Schmeil, 1898 (p.150, spp. Key); Wheeler, 1901 (p.182); Sars,1903 (p.148); Esterly, 1905 (p.203); van Breemen,1908 a (p.155, spp. Key); A. Scott, 1909 (p.186); Steuer,1915 a (p.392, Rev.); 1923 (p.5); Esterly, 1924 (p.102); Früchtl, 1924 b (p.57); Steuer, 1929 (p.497); Gurney, 1931 a (p.216); Wilson, 1932 a (p.159, spp. Key); Sewell, 1932 (p.391); Rose, 1933 a (p.267, spp. Key); Mori, 1937 (1964) (p.100, spp. Key); Dakin & Colefax, 1940 (p.105, spp. Key); Farran, 1948 (n°12, p.3); Davis,1949 (p.64); Brodsky, 1950 (1967) (p.418, spp. Key); Carvalho, 1952 a (p.150); Tanaka, 1965 (p.386); Bowman, 1965 (p.149, Rem.: rostral filaments ); Gonzalez & Bowman, 1965 (p.255); Ramirez, 1966 (p.18); M.S. Wilson, 1966 (p.109, Rem.); Subbaraju, 1967 (p.18, Table I, fig. 1, developmental time vs latitude); Owre & Foyo, 1967 (p.100, spp. Key); Björnberg, 1972 (p.73-78: Nauplius); Sheldon & al., 1972 (p.327, fig. 13: size particle vs production's rate); Bradford, 1976 (p.163 : Redef.); Razouls, 1982 (p.597); Gardner & Szabo, 1982 (p.413); Zheng Zhong & al. , 1984 (1989) (p.257, spp. Key); Mauchline, 1988 (p.715); Huys & Boxshall, 1991 (p.337, 340); Yoo & al., 1991 (Rev., p.255); Ferrari, 1992 (p.392, tab.3); Razouls, 1993 (p.307); Madhupratap & Haridas, 1994 (p.74, Rem. Subgenera); Chihara & Murano, 1997 (p.669, Subgenera & spp. Keys); Mauchline, 1998 (p.95); Bradford-Grieve, 1999 (n°181, p.3, spp. Key); Bradford-Grieve & al., 1999 (p.961, 962: spp. Key); Bradford-Grieve, 1999 b (p.214, Def., Rem.); Boxshall & Halsey, 2004 (p.52); Mulyadi, 2004 (p.137, Def., spp. Key in Indonesian waters); Vives & Shmeleva, 2007 (p.394, spp. Key) | | Rem.: | In a genus revision, Steuer in 1915 establishes two subdivisions based on the presence/absence of rostral filaments (Arostratae et Rostratae), but later Bowman (1965) when studying a population of A. liljeborgi demonstrates that the rostral filaments can be present or absent. The environmental conditions, notably the salinity, could be responsable for this fact.
It is probable that other morphological characters show a certain variability like the spinulation of the abdomen, the structure of the P5. Various anomalies have been shown by Brylinski (1984, p. 961) .
The genus divides currently into 6 sub-genera: Acartia (Planktacartia ), Acanthacartia , Acartiura , Euacartia , Hypoacartia , Odontacartia, contested by Barthélémy (1999, p.868; 1999 a, p.59, scheme 9) after analysis of the genital structures. Among the majority of species, we can distinguish two main groups, based on the external morphology of the genital area on one hand, and the seminal duct morphology on the other. The first group includes ten species, viz., A. clausi, A. hudsonica, A. omorii, A. longiremis of the subgenus Acartiura, A. bifilosa, A. chilkaensis, A. italica, A. levequei, A. tonsa of the genus Acanthacartia, and A. lilljeborgi of the subgenus Odontacartia, exhibing a compact external area, with well-defined structure for each of them. All exhibit a very homogeneous configuration of their internal area with seminal ducts in characterisytic loop-like form. According to Steuer (1923), this loop is visible in ventral view after clearing the specimens. The duct seems less voluminous only in the subgenus Acanthacartia species. The similitude in the organization of the genital structures af all these species confirms the closerelationship between the subgenera Acartiura and Acanthacartia. Thus, the morphology of the genital complex does not justify the distinction of two subgenera. The second group comprises A. danae, A. negligens of the subgenus Acartia, A. amboinensis, A. japonica of the subgenus Odontacartia and A. southwelli of the subgenus Euacartia; these species offer a fragmented external area with ventrolateral genital slits, and seminal ducts, simple bend-shaped opening in the proximal zone of the egg-laying ducts, as in the precedent group. Therefore, the genital anatomy does not justify the distinction between the different subgenera.
Total: 62 spp. + 2 undet. + 3 n. nudum.
Diagnosis from Bradford-Grieve (1999, p.3): As for the family, with the following additional characters.
- Anal somite is without an anal operculum, as anus opens between last two urosomites into a dorsal grove on anal somite.
- Caudal rami short, separated from anal somite.
- A2 basis fused with endopodal segment 1, which is long and slender and bears 9 setae; exopod shorter than endopodal segment 1.
- P1 exopodal segment 1 and 2 each with a long slender outer distal spine, and with 2 spines on exopodal segment 3.
- Female P5 3-segmented , uniramous, with last segment modified into a long, slender spine.
- Male P5 larger on right, exopodal segment 2 with a large inner lobe, and exopodal segment 3 in the form of a clasper. | | Remarks on dimensions and sex ratio: | | Females: Mean = 1.0966 mm (n = 113, SD = 0.362). Males: Mean = 0.992 mm. (n = 102; SD = 0.271). The size ratio Male: Female = 0.9046. | | | | | Acanthacartia Steuer, 1915 | | Ref.: | Steuer, 1923 (p.21, 55); Gurney, 1931 a (p.217); Rose, 1933 a (p.268); Bradford, 1976 (p.164); Chihara & Murano, 1997 (p.670); Bradford-Grieve, 1999 (n° 181, p.5); Barthélémy, 1999 a (p.32); Bradford-Grieve, 1999 b (p.214, Def., Rem.) | | Rem.: | Subgenus of Acartia defined by Steuer, 1915 a (p.396)
Type: Acartia pietschmanni Pesta, 1911. Total: 18 spp.
The mean female size is 1.146 mm (n = 34, SD = 0.4366) and for male 1.022 mm (n = 32, SD = 0.3475). The size ratio male female is 0.8918.
Diagnosis from Bradford-Grieve (1999, p.5): As for the family and generic definition, with the following additional characters.
- Rostral filaments usually present.
- Ovaries fused.
- A1 segments mostly unspined.
- Posterior prosomal borders rounded and either naked or armed with spines, sometimes large and acute.
- Spermathecal canal appears to be looped only in ventral view.
- P5 as in Acartiura although heavy spine on male left exopodal segments 2+3 is not simple spine found in Acartiura but may have one or more accessory spines arising from its base or be expanded .
- Coastal species.
| | | | Acartia Dana, 1846 (part.) | | Syn.: | Planktacartia Steuer, 1915 a (p.397); Steuer, 1923 (p.34,55);
Gurney, 1931 a (p.217); Rose, 1933 a (p.268)
| | Ref.: | M.S. Wilson, 1966 (p.109, Rem.); Bradford, 1976 (p.163); Chihara & Murano, 1997 (p.669); Bradford-Grieve, 1999 (n°181, p.6); Barthélémy, 1999 a (p.32); Bradford-Grieve, 1999 b (p.218, Def., Rem.) | | Rem.: | Type species: Acartia negligens Dana, 1849. Total: 5 spp.
The mean female size is 1.381 mm (n = 7, SD = 0.5906). The mean size male is 1.025 mm (n = 4, SD = 0.5939). The size ratio male:female = 0.901
Diagnosis from Bradford-Grieve (1999, p.6): As for the genus, with the following additional characters.
- Rostral filaments present.
- Ovaries fused.
- Posterior prosomal borders rounded or drawn out into a point.
- Female P5 coxopodite with long plumose seta, terminal spine denticulate.
- Right male P5 exopodal segment 1 with a distal appendage.
- Indopacific and Atlantic species. Oceanic in all tropical and sudtropical seas. | | | | Acartiura Steuer, 1915 | | Ref.: | Steuer, 1923 (p.5); Gurney, 1931 a (p.217, 220); Rose, 1933 a (p.267); Bradford, 1976 (p.159, Rev., spp. Key); Chihara & Murano, 1997 (p.669, clé spp.); Bradford-Grieve, 1999 (n°181, p.3); Barthélémy, 1999 a (p.31); Bradford-Grieve, 1999 b (p.221, Def., Rem.) | | Rem.: | Subgenus of Acartia defined by Steuer, 1915 a (p.394); 1923 (p.5)
Type: Acartia clausii Giesbrecht,1889. Total: 15 spp. + 2 undetemined.
Diagnosis from Bradford-Grieve (1999, p.3): As for the genus, with the following additional characters.
- Ovaries fused.
- Posteriorlateral corners of prosome rounded but may bear spines.
- Rostral filaments absent.
- Caudal rami short and slightly asymmetrical, with right ramus longest.
- Female genital double-somite with spermathecal canal looped when viewed both laterally and ventrally, and genital apertures close together on ventral surface.
- Female P5 has a smooth terminal spine bearing some distal hairs on both sides and with an evenly bulbous base, endopod absent.
- Male P5 has right basis with 2 inner ridges and an outer distal plumose seta, exopodal segment 1 (not in A. discaudata) has a distal inner lobe and a proximal inner spine, exopodal segment 2 has its inner lobe bearing 1-3 spines, exopodal segment 3 has a terminal spine, and inner edge spine and at least 3 transverse rows of outer spines. Left basis has an inner proximal ridge, postrior surface spines, and an outer distal plumose seta; exopodal segment 1 usually has some spinules; the exopodal segments 2+3 is shaped like a hand with 2 lateral distal opposing spines, anterior spine much heavier and thicker-walled than posterior spine, but both are simple; patches of spinules and hairs are proximal to anterior spine;
- Mainly temperate to cold species in both hemispheres (Bradford, 1976).
The mean female size is 0.9628 mm (n = 37, SD = 0.2131), and for male 0.8916 mm (n = 35, SD = 0.1850). The size ratio male:female is 0.926.
| | | | Euacartia Steuer, 1915 | | Ref.: | Gurney, 1931 a (p.217); Rose, 1933 a (p.268); Bradford, 1976 (p.176); Madhupratap & Haridas, 1994 (p.74); Barthélémy, 1999 a (p.33, Rem.); Bradford-Grieve, 1999 b (p.226, Def., Rem.: p.214); Soh & al., 2013 (p.718, Table 1, 2, fig.9: geographical distribution) | | Rem.: | Subgenus of Acartia defined by Steuer, 1915 a (p.394).
Type: Acartia southwelli Sewell, 1914. Total 3 spp.
The mean female size is 0.8326 mm (n = 6, SD = 0.0660), and for male 0.7516 mm (n = 6, SD = 0.0649). The size ratio male:female is 0.903.
Diagnosis from Bradford-Grieve (1999 b, p.226): As for the family with the following additional characters/
- Rostral filaments present.
- Posterior metasomal borders rounded.
- Female P5 basis long with a short outer edge seta.
See remarks in Odontacartia. | | | | Hypoacartia Steuer, 1915 | | Ref.: | Steuer, 1923 (p.14,54); Gurney, 1931 a (p.217); Rose, 1933 a (p.268); Bradford, 1976 (p.163); Bradford-Grieve, 1999 (n°181, p.6); Bradford-Grieve, 1999 b (p.226, Def.) | | Rem.: | Subgenus of Acartia defined by Steuer, 1915 a (p.396).
Type: Acartia macropus Cleve,1901. Total: 2 spp.
The mean female size is 1.207 mm (n = 3, SD = 0.1716), and for male 1.230 mm (n = 3, SD = 0.05). The size ratio male:female is 1.019
Diagnosis after Bradford-Grieve (1999, p.6): As for the family with the following additional characters.
- Rostral filaments present.
- Ovaries paired.
- Female posterior prosomal borders with asymmetrical lateral extensions.
- Female P5 inner border with a broad, dentate terminal spine.
- Male P5 superficially similar to that of A. discaudata.
- Coastal species.
| | | | Odontacartia Steuer, 1915 | | Ref.: | Steuer, 1923 (p.26, 55, 2 groups: 'erythraea' & 'centrura'); Gurney, 1931 a (p.217); Rose, 1933 a (p.268); Bradford, 1976 (p.163); Ueda, 1986 (Rem., p.17,18); Chihara & Murano, 1997 (p.671); Barthélémy, 1999 a (p.33); Bradford-Grieve, 1999 b (p.226, Def., Rem.); Srinui & al., 2019 (p.79: 2 species-groups: centrura and erythraea, p.89: females and males species key); Lee S. & al., 2019 (p.85: Key of species; p.82: phylogeny; Rem.). | | Rem.: | Subgenus of Acartia defined by Steuer, 1915 a (p.396).
Type : Acartia lilljeborgi Giesbrecht,1889. Total: 14 spp.
The mean female size is 1.305 mm (n = 22, SD = 0.1770), and for male 1.1755 mm (n = 20, SD = 0.1444). The size ratio male:female is 0.901.
Diagnosis from Bradford-Grieve (1999 b, p.226): As for the family and generic definitions with the following additional characters.
- Rostral filaments present.
- Posterior metasomal borders drawn out into lateral spines, likewise the posterior borders of the urosome segments, especially in the male (except A. lilljeborgi).
For Srinui & al. (2019, p.79) Acartia (O.) lilljeborgi is regarded as an intermediate type between the centrura and erythraea species groups (Steuer, 1923; Ueda, 1986).
The centrura species group is defined as: in the female the genital double-somite with a pair of large processes; first A1 segments without a large spine; relatively long caudal rami; exopod of P5 with a knob located or extending to midlength. In the male: 3rd and 4th urosomites with a dorsal pair of large acure processes; 1st exopodal segment of left P5 without an outer spine.
For Lee S. & al. (2019, p.82) Odontacartia can be divided into two groups based on the presence of a spine on the 1st segment of A1. Species with this spine include A. amboinensis, A. australis, A. bispinosa, A. erythraea, A. japonica, A. lilljeborji. Species lacking spine include A. bowmani, A. centrura, A. mertoni, A. ohtsukai, A. pacifica, A. spinicauda, A. nadiensis. The length ratio of the outer seta/terminal spine of the female P5 is the most diagnostic morphological feature in Odontoacartia species. However, this character is also used to determine the subgenus Euacartia (see Soh & al., 2013). This confusion between subgenus systems has been documented (Madhupratap & Haridas, 1994). Barthélémy (1999) compared female genital structure of 25 species of Acartiidae and concluded that there is no support for the subdivision of Acartia into subgenera. | | | | Planktacartia Steuer, 1915 | | Rem.: | Subgenus of Acartia according to Steuer, 1915 a (p.397). M. S. Wilson, 1966 (p.109) considers the maintenance of this subgenus to be incorrect. It should be denominated Acartia in conformity with the International Code of Nomenclature.
Type: Acartia negligens . Cf. Acartia | | | | (2) Acartiella Sewell, 1914 | |
| | Syn.: | S.G: Acartiella Steuer, 1915 a (p.394); 1923 (p.12, 54); Rose, 1933 a (p.268); Wellershaus, 1969 (p.268, spp. Key) | | Ref.: | Sewell, 1914 a (p.245); 1919 (p.2, 18, spp. Key); Gurney, 1931 a (p.215, 216, 217); Sewell, 1932 (p.392); Bradford, 1976 a (p.162); Zheng Zhong & al., 1984 (1989) (p.259); Barthélémy, 1999 (p.868, Rem.); 1999 a (p.59, Rem.); Bradford-Grieve, 1999 b (p.227, Def.); Boxshall & Halsey, 2004 (p.52) | | Rem.: | Genus defined by Sewell (1914) with for species type: Acartia kempi Sewell, 1914. Considered by various authors as a sub-genus or a new family. According to Sewell (1914) the female P5 ''possesses both an exopodite and an endopodite, and in the male the right leg forms a well-developed clasping apparatus: in both species the rostrum is absent.
For Barthélémy (1999 a, p. 59) this genus has to constitute to him only a family. The species present a genital area of diaptomoid type deprived by opercule and by receptacle seminal, which distinguishes them from all other species of the family of Acartiidae possessing a genital model of area characterized well by the constant presence of receptacles and pores copulators neighboring to gonopores. We can also observe the other morphological characters: the length of the branches of the caudal rami, the shape flattened by the antennula, the biramous P5. These characters differentiate all the species of the genus of the other Acartiidae, with the exception of Acartiella kempi, only species of the genus presenting an antennule similar to that of Acartia (Sewell, on 1918).
According to Barthélémy (1999, p.868) the morphology of the external genital area and the absence of seminal receptacles very clearly isolate species of the genus Acartiella Sewell (1914), the validity of which is thus confirmed. Bradford (1976) pointed out that Acartiella should remain in the Acartiidae family especially because the exopods of P2-P4 are devoid of articulated outer edge spines, as in the other Acartiidae.
Total: 11 spp.
The mean female size is 1.166 mm (n = 18), and for male 1.062 mm (n = 19). The size ratio male/female is 0.911. | | Remarks on dimensions and sex ratio: | | The mean female size is 1.166 mm. (n = 18, SD = 0.247) and the mean male size is 1.062 mm. (n = 19, SD = 0.223). The size ratio male: female is 0.911. | | | | (3) Paracartia T. Scott, 1894 | |
| | Syn.: | Paracartia T. Scott, 1894 b (p.68); Giesbrecht, 1897 b (p.254); Steuer, 1915 a (p.394); 1923 (p.15, 54); Rose, 1933 a (p.268) | | Ref.: | Sars, 1904 b (p.3); 1919 (1921) (p.15); Gurney, 1931 a (p.217); Sewell, 1948 (p.378, Rem.); Gonzalez & Bowman, 1965 (p.255); Bradford, 1976 (p.162, 163); Dussart, 1982 (p.20); Razouls,1982 (p.619); Ferrari, 1992 (p.392, tab.3); Razouls, 1993 (p.307); Bradford-Grieve, 1999 (n°181, p.14, spp. Key); Barthélémy, 1999 (p.867, 869: Rem.); 1999 a (p.34, 59, Rem.); Bradford-Grieve, 1999 b (p.227, Def.); Boxshall & Halsey, 2004 (p.52); Vives & Shmeleva, 2007 (p.433, spp. Key) | | Rem.: | Considered as a subgenus by T. Scott, Steuer, then later as a genus by Sars (1904b).
For Barthélémy (1999 a, p. 59) the organization of the external genital area , separated, in very lateral side and posterior genital splits, could constitute an important characteristic of the genus.
Type: Acartia dubia T. Scott,1894. Total: 5 spp.
Diagnosis from Bradford-Grieve (1999, p.14):
- Rostral filaments present.
- Sexual dimorphism strongly marked.
- Ovaries paired.
- Last prosomite of female usually strongly spread into wing-shaped extensions on each side, that of the male simple with lateral parts not expanded.
- Urosome of female comparatively short, 3 free somites with genital double-somite expanded laterally, may be asymmetrical, genital pores laterally placed on somite.
- Caudal rami broad and 1 of apical setae may or may not be transformed into a thickish spine.
- Urosome of male slender, of 5 somites, with caudal rami of normal shape.
- Female A1 as in Acartia.
- Right geniculate A1 of male may be expanded.
-A2 mouth parts, and swimming legs similar to those of Acartia.
- Female P5 with both basis fused, terminal spine, if present, is heavily built, and lateral seta is usually much shorter than terminal spine.
- Male P5 powerfully developed; right leg much larger and terminates in a very long slender incurved claw, basis with a conspicuous inner projection which is sometimes very large; left exopodal segments 2 + 3 usually wider than long, bearing at least 2 spines which oppose each other to make segment resemble a chela.
- The spermatophore, fixed to genital-double-somite of female is accompanied by a complicated thin plate which may curve around female genital double-somite. | | Remarks on dimensions and sex ratio: | | The mean female size is 1.106 mm. (n = 9, SD = 0.244) and the mean male size is 1.030 mm. (n = 9, SD = 0.204). The size ratio male: female is 0.931. | | | | (4) Paralabidocera Wolfenden, 1908 | |
| | Ref.: | Wolfenden, 1908 (p.26); Vervoort, 1951 (p.148, Rem.); Bradford, 1976 (p.163); Brodsky & Zvereva, 1976 a (p.233-34, Rem.); Razouls,1982 (p.620); 1993 (p.307); Mauchline, 1998 (p.95); Barthélémy, 1999 (p.869: Rem.); 1999 a (p.34); Bradford-Grieve, 1999 b (p.227, Def., Rem.); Boxshall & Halsey, 2004 (p.52) | | Rem.: | Type: Paracartia antarctica Thompson,1898. Total: 3 spp.
For Barthélémy (1999, p.869) the sole examined species presents a fragmented area with two lateral genital slits, but internally there is no seminal duct and the receptacle seems to open distally in the egg-laying duct. The presence of paired genital orifices situated on both side of the genital segment is a constant character of the genus. The three species of the genus therefore present genital structures clearly differing from those of the Pontellidae. Thus, in spite of the resemblance with the genus Labidocera (Pontellidae), the comparison with the genital structures in Pontellidae (Barthélémy & al., 1998) confirms that the genus Paralabidocera must be referred to Acartiidae.
Diagnosis from Bradford-Grieve (1999 b, p.227): As for the family definition with the following additional charaters.
- 2 rostral filaments present.
- Posterior metasomal corners rounded.
- Female genital segment swollen and may be asymmetrical.
- Anal operculum absent.
- Female P5 with fused 1st basal segments, an endopod, and a bifurcate 1-segmented exopod.
- Male P5 generally as in other Acartiidae but much more powerfully built.
- The spermatophore has a larger coupler which wraps around both sides of the female genital segment. | | Remarks on dimensions and sex ratio: | | The mean female size is 1.718 mm. (n = 6, SD = 0.257), and the mean male size is 1.510 mm. (n = 6, SD = 0.156). The size ratio male: female is 0.879. | | | | (5) Pteriacartia Belmonte, 1998 | |
| | Syn.: | Acartia josephinae Crisafi, 1974 (p.6) | | Ref.: | Belmonte, 1998 (p.360); Bradford-Grieve, 1999 (n°181, p.15); Barthélémy, 1999 a (p.34); Boxshall & Halsey, 2004 (p.52); Vives & Shmeleva, 2007 (p.436) | | Rem.: | type: Acartia josephinae. Total: 1 sp.
Diagnosis from Bradford-Grieve (1999, p.15), after Belmonte, 1958 :
- Rostral filaments present.
- Sexual dimorphism marked.
- Last prosomite of female and male rounded.
- Urosome of female comparatively short, of 3 free somites with genital double-somite asymmetrical, longer than half the urosome, genital pores paired, laterally placed, withot opercula; fin-like expansion on right side of genital double-somite.
Male urosome slender, of 5 free somites, caudal rami short.
- Female A1 22-segmented.
- Right geniculate A1 of male without swollen segments.
- Female P5 reduced to small knob, each with 1 long seta.
- Male P5 right leg much larger than left leg and terminates in a long claw which is curved inwards; exopodal segment 2 of right leg with a long finger-like protrusion originating from the distal half of the segment; left leg exopodal segments 2 + 3 half-moon shaped with spines at either end.
- The spermatophore is fixed by a gelatinous 'apron' to the ventral surface of female genital double-somite. | | Remarks on dimensions and sex ratio: | | From only an species in the genus, the minimum and maximum female size is 0.796 mm and 0.865 mm, (mean: 0.8305 mm), and for male: 0.788 mm and 0.865 mm (mean: 0.8265 mm). The size ratio (male: female) is 0.995. | | | | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed July 05, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|



;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
