|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
| |
| | | |
| Pseudodiaptomidae Sars, 1902 ( Diaptomoidea ) | | Ref.: | Sars, 1902 (1903) (p.73); Gurney, 1931 a (p.84); Wright, 1937 (p.160, clé spp.); Nicholls, 1944 (p.8); Brodsky, 1950 (1967) (p.82, 322); Gonzalez & Bowman, 1965 (p.250); Björnberg, 1972 (p.49); Andronov, 1974 a (p.1005); Kiefer, 1978 d (p.55, 57); Madhupratap & Haridas, 1978 (p.257, Rem.); Bowman & Abele, 1982 (p.9); Razouls, 1982 (p.435); Dussart & Defaye, 1983 (p.29); Brodsky & al., 1983 (p.143, 146); Grindley, 1984 (p.217, Rem.: 5 groups, biogeography); Sazhina, 1985 (p.63, 116, N); Zheng Zhong & al.,1984 (1989) (p.245, Rem.); Walter, 1986 (p.129); 1986 a (p.504); Dussart, 1989 (p.6); Grindley, 1990 (p.237, Biogeography); 1990 a (p.237); Huys & Boxshall, 1991 (p.419); Razouls, 1993 (p.308); Chihara & Murano, 1997 (p.893); Bradford-Grieve & al., 1999 (p.884, 902, 904, 952); Bradford-Grieve,1999 b (p.148, Def., Rem.); Ohtsuka & Huys, 2001 (p.461); Boxshall & Halsey, 2004 (p.13; 49; 172: Def.; p.174: Genera key); Mulyadi, 2004 (p.152); Laakmann & al., 2019 (p.330, fig. 2, 3, phylogenetic relationships)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm  | | Rem.: | 3 G.: Archidiaptomus, Calanipeda, Pseudodiaptomus. These brackish and freshwater forms, sometimes littoral, are not yet taken completely into consideration (incomplete references).
Key to genera after Boxshall & Halsey (2004, p.174) :
1 - Female P5 biramous with well developed endopod; male right A1 with 4 segments distal to geniculation ....... Archidiaptomus arrorus.
1' - Female P5 uniramous, endopod absent; male right A1 with at most 3 segments distal to geniculation ..... 2.
2 - Female A1 25-segmented; A2 exopod 6-segmented .........Calanipeda aquaedulcis.
2' - Female A1 20 to 23-segmented; A2 exopod 4-segmented .......... Pseudodiaptomus.
Definition from Bradford-Grieve (1999 b, p.148) :
Female :
- Cephalosome and pediger 1 fused or separate, pedigers 4 and 5 fused or separate.
- Usually eye composed of a pigmented spot surrounded by a refractible lens.
- Posterior metasomal corners may be rounded or variously decorated with large posterior spines or rows of spinules.
- Urosome 2-4-segmented, may be asymmetrical.
- Genital segment variable (symmetrical or asymmetrical, ornamented with spines and /or spinules, or undecorated).
- Caudal rami tend to be elongate and may be asymmetrical.
- A1 symmetrical, usually 22-segmented (24-segmented in Archidiaptomus, 21-segmented in the hyalinus group).
- A2 exopod slightly longer than endopod, endopod with fusion between segments and with basis.
- Md blade with numerous small teeth.
- Mx1 usually with inner lobe 1 (arthrite) with 15 spines and setae ; inner lobes 2 and 3 wiyj 3 setae each ; basis with 5 setae ; endopodal segments 1-3with 4, 4, 5 setae respectively ; exopod with 10 setae ; outer lobe 1 with 10 setae.
- Mx2 inner lobes 1-5 with 4, 3, 3, 3, 3 setae respectively.
- Mxp squat, coxa with 2, 3, 4 setae, one of the terminal group enlarged into a spine ; basis bordered by spinules and 3 setae ; endopod 4-segmented carrying 2-4 characteristic setiform bristles (2 on segment 1 and 1 each on segments 2 and 3)
- Swimming legs biramous, with 3-segmented rami ; basipods 1 and 2 (coxa and basis) often decorated with spinules on anterior, posterior and lateral surfaces, also sometimes on anterior surfaces of endopods and exopods.
Male :
- Urosome 5-segmented.
- A1 asymmetical ; left 22- or 24-segmented ; right with 20-21 segments, geniculate between segments 18 and 19 ; terminal part beyond the joint can have from 2 to 4 segments.
- Mouthparts similar to those of female | 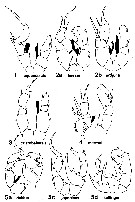 issued from : J.R. Grindley in Crustaceana, 1984, supplt 7. [p.219, Fig.1]. Examples of the five groups of Pseudodiaptomidae distinguished by the arrangement of the endopodites of male P5. Endpodites dermarcated in black in these diagram. Nota : For Grindley (1984, p.218) there are several different groups of species which tend to constitute morphologically and zoogeographically distinct groups. The most important charcters distinguishing the species are the P5, and in particular those of males ; in particular the arrangement of the endopods of the P5 in the males is different for each of several groups and comparable whitin each group. All the species found round the coast of Africa form a group characterized by the possession of both left and right endopods in the P5 of the male (Grindley, 1963). Certain oriental species possess both endopods but they be separated on the possession of a Y-shaped right endopod in the male P5. Group 1 : Both endopodites present, of which neither is greatly reduced ( Calanipeda, Archidiaptomus ; Mediterranean, Ponto-Caspian region, and Cochin). Group 2 : Both endopodites present of which the right is greatly reduced. (Africa, Asia, Australia). Group 3 : Only the left endopodite present (North and South America). Group 4 : Only the right endopodite present. (Asia and Australia). Group 5 : A long curved medial projection in place of the left endopodite on the second left nasal segment ; right endopodite reduced or absent. (Asia and Australia). Other features appear to agree, for example, the form of the P5 females (used by Sewell, 1924, in his attempt at grouping, fit the same pattern. The simplest and apparently most primitive form of the endopodite appears to be exhibited by Calanipeda aquaedulcis and Archidiaptomus arroorus. In addition to its characteristic endopodites, C. aquaedulcis is unique among Pseudodiaptomidae in having A1 with 25 segments (primitive condition in calanoid copepods). The segmentation of the male geniculate A1 is reduced in Calanipeda, but there are more end segments beyond the articulation than in any other pseudodiaptomid. Archidiaptomus aroorus possesses a number of primitive characters and strong affinities to the family Diaptomidae ; it has been placed in this new genus because of a number of peculiar features including the fully developes left and right endopods in the male P5 ; A1 is 24-segmented which approaches the primitive condition of the Calanoida ; exopod 2-segmented of the female P5 is close to that of the typical Pseudodiaptomidae but it is unique in maintening fully developed endopods of an unusually spinous form (this appears to be a primitive or generalized condition placing this genus very close to the Diaptomidae). |
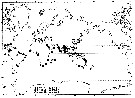 issued from : J.R. Grindley in Crustaceana, 1984, supplt 7. [p.224, Fig.2]. Distribution of the five groups of Pseudodiaptomidae. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.172]. Armature formula of swimming legs P1 to P4. Nota: Spine and setal formula sometimes reduced. Nota : Female P5 biramous with 1-segmented endopods in Archidiaptomus, uniramous in other genera ; uniramous leg typically 5-segmented, symmetrical, primitively joined by distinct intercoxal slerite or with coxae fused ; comprising coxa, basis and 3-segmented exopod. Coxa unarmed. Basis with outer seta. 1st exopodal segment typically elongate ; 1st and 2 nd exopodal segments each with outer spine, 2 nd produced into process at inner distal angle. 3rd exopodal segment small, fused to apical element and with small inner process proximally. - Male P5 asymmetrical, primitively biramous, uniramous due to loss of endopod in some species ; separate coxa and basis present on both sides ; right leg with 3-segmented exopod, endopod 1-segmented, often bifid ; endopod sometimes absent. 3rd exopodal segment typically curved, sometimes claw-like, armed with 1 large seta and 1 or 2 setules on concave margin. Left leg with 2-segmented exopod, 1st segment with outer spine, 2 nd segment irregularly shaped, with distal spines. Endopod typically an elongate , unsegmented lobe. - Eggs contained in paired, multiseriate sacs, or in a single sac according to species. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.173]. A, Pseudodiaptomus ishigakiensis habitus female; B, habitus male; C, female P5; D, male P5. E, Pseudodiaptomus hessei female A2; F, female Md. [Nishida, 1985a: A-D; Grindley, 1963: E-F. |
 Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.148]. Spine and setae formula in swimming legs P1 to P4. P5 female uniramous or biramous ( Archidiaptomus), may be slightly asymmetrical; coxa and basis separate, basis usually with a posterolateral seta; exopod 3-segmented (terminal 2 segments fiused in Archidiaptomus. Ovisac present. P5 male uniramous or biramous, asymmetrical; exopods 2- or 3-segmented; spines , rxopod al segments, and endopods, if present, variously modified into organs which appear to be adapted for clasping. | | | | | (1) Archidiaptomus Madhupratap & Haridas, 1978 | |
| | Ref.: | Madhupratap & Haridas, 1978 (p.253); Razouls, 1982 (p.445); 1993 (p.308); Dussart & Defaye, 1983 (p.36); Grindley, 1984 (p.217, 218, 220); Walter, 1986 (p.129); Mauchline, 1998 (p.67); Bradford-Grieve,1999 b (p.148, Def.); Boxshall & Halsey, 2004 (p.174) | | Rem.: | Type: Archidiaptomus arrorus Madhupratap & Haridas, 1978. Total : 1 sp.
Brackish water on the coast of India.
After Madhupratap & Haridas (1978, p.257) this genus and species possesses many 'primitive' characters and strong affinities to the family Diaptomidae. The 2-segmented exopod of the P5 female (though somewhat peculiar in form) is typical of the Pseudodiaptomidae; but it is unique in maintaining fully developed endopods of an unusually spinous form. This appears to be a primitive or generalised condition placing this genus very close to the Diaptomidae. The exopods of the right and left P5 male are fully developed as in the Pseudodiaptomidae (in the Diaptomidae the left one is usually smaller). But it is again unusual in maintaining the primitive feature of both endopods being fully developed whereas in Pseudodiaptomus the endopods of the P5 male are either rudimentary or lacking. The urosome is 4-segmented in the female (3-segmented in the Diaptomidae) and 5-segmented in the male as in the Pseudodiaptomidae and the ornamentation of the genital segment is also somewhat pseudodiaptomid in character. The A1 has a greater number of segments than the species of thje genus Pseudodiaptomus (mostly 20 to 22, 23 in some) and in having 24 segments approaches the 25-segmented nature of the diaptomid A1, which is considered as a primitive condition in calanoid Copepoda. The right geniculate A1 of the male has 4 terminal segments as is usually found in the Diaptomidae (2 to 3 in the Pseudodiaptomidae). The P1 has a 3-segmented endopod typical of the Pseudodiaptomidae as against the 2-segmented condition in the Diaptomidae; while the unarmed nature of the outer margin of the 2nd exopodal segment of P1 is a feature shared by the species of both families, the rounded form ofv the posterior angles of the metasome and the long caudal rami of the present type-species are pseudodiaptomid characters.
Definition from Bradford-Grieve (1999 b, p.148) :
- As for the family definition.
- Head and pedigerous segments 1 and 2 fused, pedigerous segments 4 and 5 fused.
- Female urosome 4-segmented.
- Female genital segment symmetrical, with small anterolateral swellings, decorated with spinules.
- A1 24-segmented.
- P1 exopodal segment 3 without outer edge spines, endopodal segment 3 with 6 setae.
- Female P5 biramous, symmetical; endopod 1-segmented with a terminal claw-like process at the tip; exopodal segment 1 with 1 single long outer spine, exopodal segment 2 with 4 spines, terminal one the longest.
- Male P5 biramous, asymmetrical; endopods on both sides extending to middle of exopodal segment 2; left exopodal segment 1 with 1 outer distal spine and 1 longer curved inner spine, exopodal segment 2 with 1 long proximal spine and the segment terminates in 2 claw-like spines; right leg exopodal segment 2 with 1 inner spinule and 1 outer spine, segmentation between exopodal segments 2 and 3 indistinct, the latter forms a long spine with a proximal inner spinule. | | | | (2) Calanipeda Kritschagin, 1873 | |
| | Syn.: | Poppella Richard, 1888 (p.43); de Guerne & Richard, 1889 (p.149); Giesbrecht & Schmeil, 1898 (p.62) | | Ref.: | Dussart, 1967 a (p.85); Grindley, 1984 (p.217); Razouls, 1982 (p.435); 1993 (p.308); Walter, 1986 (p.129); Dussart, 1989 (p.12); Bradford-Grieve,1999 b (p.148, Déf.); Boxshall & Halsey, 2004 (p.174) | | Rem.: | Type: Calanipeda aquaedulcis Krichagin, 1873. Total: 1 sp.
Euryhaline, in estuaries around the Mediterranean, Black Sea and in the Caspian Sea.
Definition from Bradford-Gieve (1999 b, p.148) :
- As for the family definition with the following additional characters after Dussart (1967).
- Head and pedigerous segment 1 separate, pedigerous segments 4 and 5 fused.
- Posterior border of prosome rounded.
Urosome 4-segmented in female.
- Urosome male 5-segmented.
- Caudal rami 6-7 times as long as wide, inner edges hairy, with 5 well-developed setae.
Female genital segment swollen anteriorly with left side extended into a curved hook directed posteriorly and carrying a sensory spine.
- A1 25-segmented, segment 25 very small extending to posterior border of urosomal segment 2.
- Exopod of A2 6-segmented.
- Male A1 prehensile on right without either a hook nor hyaline lamella on three last segments.
- P1 to P4 with 3-segmented exopods and endopods.
- Female P5 slightly asymmetrical, without endopods; exopod 3-segmented, exopodal segment 2 with 1 strong inner spine, a strong, distal, spiniform extension, and at its base 1 small strong spine; exopodal segment 3 in the form of a powerful hook, curved inwards with 1 strong internal spine and a small spinifotm extension at its base.
- Male P5 basipod 2 large on right with 1 short seta at postero-internal angle; exopod 3-segmented, exopodal segment 1 slightly elongate with a moderate spine on postero-external angle; the two last segments of exopod form a powerfull hook; exopodal segment 2 strongly curved towards exterior and with a long extension and with 1 short slender marginal seta at midlength; exopodal segment 3 thin, long, and pointed terminally with 1 fine seta on its external border; endopod 1-segmented extending to distal border of exopodal segment 1 and armed with small spines and some terminal hairs. Left leg much shorter than right; exopod 2-segmented ; exopodal segment 1 with an elongate spiniform extension at antero-external angle reaching the distal end of exopodal segment 2 which is large, conical, and terminated by 2 subequal points; endopod 1-segmented, enlarged distally, reaching the base of exopodal segment 2, carrying at its extremity small unequal spines. | | | | | (3) Pseudodiaptomus Herrick, 1884 | |
| | Syn.: | Weismannella Dahl,1894 c (p.19);
T. Scott, 1894 b (p.39);
Schmackeria Mrazek,1894; Mazellina Rose, 1957 (p.235); Walter & al., 2006 (p.215-216, Rem.) | | Ref.: | Giesbrecht & Schmeil, 1898 (p.63); A. Scott, 1909 (p.116); Früchtl, 1924 b (p.47); Sewell, 1924 (p.784, Rem.); Gurney, 1931 a (p.20); Sewell, 1932 (p.233, Rem.); Wilson, 1932 a (p.101); Wright, 1937 a (p.160, clé spp.M); Dakin & Colefax, 1940 (p.89); Nicholls, 1944 (p.8); Brodsky, 1950 (1967) (p.322); Carvalho, 1952 a (p.145); Ummerkutty, 1960 (p.184: Rem.); Kasturirangan, 1963 (p.35); Tanaka, 1963 (p.12); Johnson, 1964 (p.33); Gonzalez & Bowman, 1965 (p.250); Wellershaus, 1969 (p.254); Andronov, 1974 a (p.1005); Pillai, 1976 (1980) (p.243, Rem., groups Key, Biogeo); Razouls, 1982 (p.436); Ranga Reddy & Radhakrishna, 1982 (p.255); Dussart & Defaye, 1983 (p.29); Zheng Zhong & al., 1984 (1989) (p.246); Grindley, 1984 (p.217); Walter, 1984 (p.369); 1986 (p.131, 162 : groups & sub-groups Key); 1986 a (p.502); 1987 (p.363, 365: clé spp., Rem., p.366, Biogeo.); Mauchline, 1987 (p.712); Dussart, 1989 (p.6, clé spp.); Walter, 1989 (p.590, spp. Key, Rem.); Jacoby & Greenwood, 1991 (p.405, species coexisting); Razouls, 1993 (p.308); Chihara & Murano, 1997 (p.893); Mauchline, 1998 (p.67); Bradford-Grieve & al., 1999 (p.884, 953: spp. Key); Bradford-Grieve,1999 b (p.149, Déf., Rem.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.174); Mulyadi, 2004 (p.152, spp. Key in Indonesian waters, Rem.: p.162); Walter & al., 2006 (p.203, 212: key of the 14 species known from the Philippines, Rem.: Revised species groups, key of species group and sub-groups) | | Rem.: | Vyshkvartzeva (1980: pers. comm.) draws attention to the fact that considering the genus Mazellina as a synonym of Pseudodiaptomus is an opinion which does not carry away conviction. For Vervoort (1965, p.97) there is no doubt on the identity of the two genera (M. galletti seems very close to P. dauglishi). Type: Pseudodiaptomus pelagicus Herrick, 1884. Coastal forms, brackish, freshwater; epibenthic in daytime. Total: 80 spp. + 2 varieties + 2 unident.
According to Walter & al., 2006 (p.203, 213, Table 2) among the 77 currently recognized species, the mouthparts and swimming legs are almost identical in shape, segmentation, spination, and/or spinulation patterns. The female genital double-somite is of particular interest (Soh & al, 2001; Walter & al., 2002 [see also Barthélémy, 1999 a]) especially regarding the ventral genital flaps and egg sac number. A1 segmentation has 3 basic patterns within this genus. The P5 females is typically symmetrical and of limited value to separate the species; the male P5 typically provides the most reliable morphological characters used for species determination and species group placement, notably the presence and /or absence of the left and /or right endopods easily indicates to which group a species belongs.
Definition from Bradford-Grieve (1999 b, p.149) :
- As in the family definition.
- Head and pedigerous segment 1 fused or separate, pedigerous segments 4 and 5 fused.
- Posterior prosomal corners often extended into points.
- Female urosome 2-4-segmented.
- Female genital segment variously ornamented, may be asymmetrical.
- Female P5 uniramous with 3-segmented exopods; in posterior view - basipod 2 with 1 large and 1 small surface seta; exopod segment 1 with a distal outer spine and 2 surface setae; exopod segment 2 with 1medial setz produced into a spiniform process which is plumose or spinulose along both margins, and 1 outer small naked ot plumose spine; exopod segment 3 spiniform, distally produced, usually equal in length or longer than exopod segment 2 spiniform process, with both margins hairy, also with a proximomedial spiniform process.
- Male P5 uniramous or biramous. Posterior view - right leg basipod 1 with a subapical spinule row; basipod 2with 1 large plumose seta and at least 1 small surface seta; exopod segments 1-3 each with at least 1 small surface seta,exopod segment 1 with 1 outer spine, exopod segment 3 proximally thickened with a medial basal swelling or process, concavely produced, and distal half of medial margin hairy, with 1-2 setae; left leg basipods 1 and 2 as on right leg; with or without an endopod; exopod segment 1 with at least 1 surface seta and variably shaped; exopod segment 2 with several surface setae, outer spine near midlength, and typically with 1 terminal spine; in anterior view: right leg basipod 1 with a hair or spinule row; basipod 2 possesses lateral spinule row that continues onto surface at midlength, and usually with an endopod; exopod segment 2 with variably shaped outer spine; left leg basipods 1 and 2 ornamented as on the right. | | Remarks on dimensions and sex ratio: | | The mean female size is 1.357 mm (n = 56; SD = 0.2776) , and the mean male size is 1.087 mm (n = 55; SD = 0.2271). The size ratio (male : female) is 0.81 (n = 64; SD = 0.0876). | | | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed March 23, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|



;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
