 List of the studied authors
List of the studied authors
Wilson, 1932 (p.38); 1942 a (p.193); Sewell, 1948 (p.325, 515); C.B. Wilson, 1950 (p.262); Yamazi, 1958 (p.153, Rem.); Heinrich, 1961 (p.92); Fagetti, 1962 (p.40); Cervigon, 1962 (p.181, tables: abundance distribution); Ganapati & Shanthakumari, 1962 (p.11, 16); Grice & Hart, 1962 (p.287, table 3); Giron-Reguer, 1963 (p.57); V.N. Greze, 1963 a (tabl.2); Grice, 1963 a (p.496); Gaudy, 1963 (p.30, Rem.); Björnberg, 1963 (p.68, Rem.); De Decker, 1964 (p.16, 22, 29, 32); De Decker, 1964 a (p.157); De Decker & Mombeck, 1964 (p.13); El-Maghraby, 1965 (p.58, Appendix); Neto & Paiva, 1966 (p.36, Table III); Furuhashi, 1966 a (p.295, vertical distribution in Kuroshio region, Table 9); Mazza, 1966 (p.73); Pavlova, 1966 (p.45); Ehrhardt, 1967 (p.742, geographic distribution, Rem.); Séguin, 1968 (p.488); Delalo, 1968 (p.138); Deevey, 1971 (p.224); Salah, 1971 (p.320); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night, Table II: %); Apostolopoulou, 1972 (p.329, 376); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Palet, 1975 (p.660); Vives & al., 1975 (p.54, tab.II, III); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Madhu Pratap & al., 1977 (p.138, Table 3: abundance vs. stations); Dessier, 1979 (p.201, 207); Vaissière & Séguin, 1980 (p.23, tab.2); Sreekumaran Nair & al., 1981 (p.493), Vives, 1982 (p.295); Kovalev & Shmeleva, 1982 (p.85); Dessier, 1983 (p.89, Tableau 1, Rem., %); Turner & Dagg, 1983 (p.17, 22); Tremblay & Anderson, 1984 (p.9); Scotto di Carlo & al., 1984 (p.1043); De Decker, 1984 (p.318, 354: chart); 1984 a (p.157); Regner, 1985 (p.11, Rem.: p.40); Binet, 1985 (p.85, tab.3); Sarkar & al., 1986 (p.178); Brinton & al., 1986 (p.228, Table 1); Renon, 1987 (tab.2); Lozano Soldevilla & al., 1988 (p.61); Dessier, 1988 (tabl.1); Böttger-Schnack & Schnack, 1989 (p.27, vertical distribution vs cyanobacteria); Böttger-Schnack, 1989 (p.33); Othman & al., 1990 (p.561, 564, Table 1); Madhupratap & Haridas, 1990 (p.305, Rem: p.317); Hirakawa & al., 1990 (tab.3); Mitra & al., 1990 (fig.3); Böttger-Schnack, 1991 (p.309); Yoo, 1991 (tab.1); McKinnon, 1991 (p.471); O'Neil & Roman, 1994 (p.235); Godhantaraman, 1994 (tab.6, 7); Kouwenberg, 1994 (tab.1); Hwang & Turner, 1995 (p.207, Table 1, 2, fig. 1, 2, 3: swimming behaviour); Webber & Roff, 1995 (tab.1); Shih & Young, 1995 (p.75); Böttger-Schnack, 1995 (p.93); 1996 (p.1088); Webber & al., 1996 (tab.1); O'Neil & al., 1996 (p.89, ingestion); Böttger-Schnack, 1997 (p.410); Suarez-Morales & Gasca, 1997 (p.1525); Park & Choi, 1997 (Appendix); Sharaf & Al-Ghais, 1997 (tab.1); Go & al., 1997 (tab.1); Noda & al., 1998 (p.55, Table 3, occurrence); Hure & Krsinic, 1998 (p.103); Padmavati & al., 1998 (p.349); Hopcroft & al., 1998 (tab.2); Gilabert & Moreno, 1998 (tab.1,2); Alvarez-Cadena & al., 1998 (t.1,2,3,4); Suarez-Morales & Gasca, 1998 a (p.113); O'Neil, 1998 (p.43, nutrition & behaviour vs
Trichodesmium substrat); Harvey & al., 1999 (p.1, 49: Appendix 5, in ballast water vessel); Siokou-Frangou, 1999 (p.478); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); El-Serehy, 1999 (p.172, Table 1, occurrence); Neumann-Leitao & al., 1999 (p.153, tab.2); Razouls & al., 2000 (p.343, Appendix); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); Fernandez-Alamo & al., 2000 (p.1139, Appendix); Lopez-Salgado & al., 2000 (tab.1); Vukanic, 2003 (p.139, tab.1); Hsieh & al., 2004 (p.398, tab.1); Rezai & al., 2004 (p.486, tab.2, 3, abundance, Rem., p.495, tab.8); Chang & Fang, 2004 (p.456, tab.1); Daly Yahia & al., 2004 (p.366, fig.4); Lan & al., 2004 (p.332, tab.1); Gallienne & al., 2004 (p.5, tab.3); Kazmi, 2004 (p.230); Choi & al., 2005 (p.710: Tab.III); Baker & al., 2005 (tabl.); Smith & Madhupratap, 2005 (p.214, tab.4,5); Berasategui & al., 2005 (p.485, tab.1); Berasategui & al., 2006 (p.485: fig.2); Rakhesh & al., 2006 (p.93, Table 2, spatial distribution); Hwang & al., 2006 (p.943, tabl. I); Dias & Araujo, 2006 (p.78, Rem., chart); Khelifi-Touhami & al., 2007 (p.327, Table 1); Hwang & al., 2007 (p.25); Eberl & Carpenter, 2007 (p.205); Jitlang & al., 2008 (p.65, Table 1); Rakhesh & al., 2008 (p.154, abundance vs stations); Tseng L.-C. & al., 2008 (p.153, fig.5, Table 2, occurrence vs geographic distribution, indicator species); Humphrey, 2008 (p.85: Appendix A); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.514, 524); Fernandes, 2008 (p.465, Tabl.2); Pagano, 2009 (p.115); C.-Y. Lee & al., 2009 (p.151, Tab.2); Miyashita & al., 2009 (p.815, Tabl.II); Tseng & al., 2009 (p.327, fig.5, feeding); Cornils & al., 2010 (p.2076, Table 3); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Dias & al., 2010 (p.230, Table 1, fig.5 a); Mazzocchi & Di Capua, 2010 (p.430); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Fazeli & al., 2010 (p.153, Table 1); W.-B. Chang & al., 2010 (p.735, Table 2, abundance); Hsiao S.H. & al., 2011 (p.475, Appendix I); Tseng L.-C. & al., 2011 (p.239, Table 3, abundance %); Hsiao & al., 2011 (p.317, Table 2, indicator of seasonal change); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); Pepin & al., 2011 (p.273, Table 2, seasonal abundance); Isari & al., 2011 (p.51, Table 2, abundance vs distribution); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Shiganova & al., 2012 (p.61, Table 4); Uysal & Shmeleva, 2012 (p.909, Table I); Salah S. & al., 2012 (p.155, Tableau 1); Jose & al., 2012 (p.20, fig.3 a,b,c: % vs monsoon); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Zizah & al., 2012 (p.79, Tableau I, Rem.: p.86); Johan & al., 2012 (2013) (p.1, Table 1); Miloslavic & al., 2012 (p.165, Table 2, transect distribution); Tseng & al., 2012 (p.621, Table 3: abundance); Lavaniegos & al., 2012 (p. 11, Appendix); Belmonte & al., 2013 (p.222, Table 2, abundance vs stations);
in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Melo Jùnior & al., 2013 (p.363, Table 2, egg production); Tseng & al., 2013 (p.507, seasonal abundance); Tseng & al., 2013 a (p.1, Table 3, abundance); Rakhesh & al., 2013 (p.7, Table 1, abundance vs stations); Jagadeesan & al., 2013 (p.27, Table 3, 5, 6, fig.11, seasonal variation); Peijnenburg & Goetze, 2013 (p.2765, genetic data); Lidvanov & al., 2013 (p.290, Table 2, % composition); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance) ; Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Zaafa & al., 2014 (p.67, Table I, occurrence); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production, Table 4: % vs. season); Nakajima & al., 2015 (p.19, Table 3: abundance); Rojas-Herrera & al., 2016 (p.40, Table 2: temporal abundance); Zakaria & al., 2016 (p.1, Table 1) ; Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Ben Ltaief & al., 2017 (p.1, Table III, Summer relative abundance); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence); El Arraj & al., 2017 (p.272, table 2, seasonal composition); Bonecker C.T. & al., 2017 (p.247, fig.10: abundance day/night vs. vertical types of mass water); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Dias & al., 2018 (p.1, Tables 2, 4, 5: vertical distribution, abundance vs. season); Palomares-Garcia & al., 2018 (p.178, Table 1: occurrence); Acha & al., 2020 (p.1, Table 3: occurrence % vs. ecoregions).


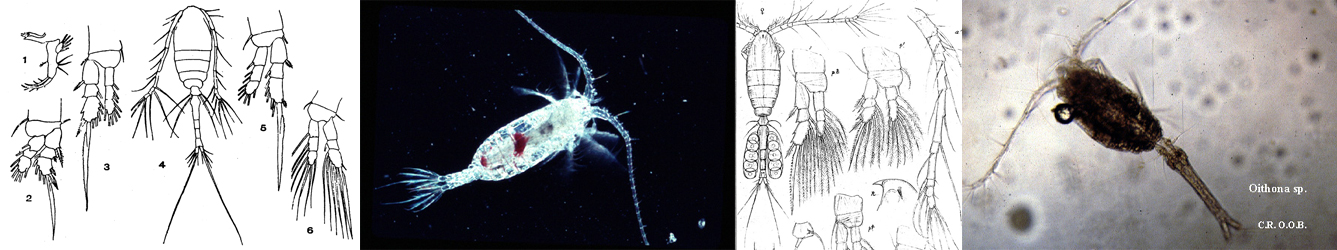

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}