 List of the studied authors
List of the studied authors
T. Scott, 1902 (p.454); Sewell, 1948 (p.367, 419, 483, 487); Conover, 1959 (p.259, Table 1, 2, respiration); M.W. Johnson, 1961 (p.311, Table 2); H. Schulz, 1961 (p.57); Marshall & Orr, 1962 (tab.3); Cannicci, 1962 (p.349, Table3); ? Shih & al., 1971 (p.29, 200); Paulmier, 1971 (p.168); Martens, 1972 (p.15, fecal pellets); Arndt & Heidecke, 1973 (p.599, 603); Eriksson, 1973 a (p.95, fig.10, area abundance distibution); Castel & Lasserre, 1977 (p.129, fig.2, 3, 7); Ciszewski & Witek, 1977 (p.449, P/B quotient); Hernroth, 1978 (p.1, Rem.: p.5); Collins & Williams, 1981 (p.273, monthly distribution-salinity); Castel & Courties, 1982 (p.417, Table II, fig.4, spatial & monthly distribution); Tremblay & Anderson, 1984 (p.3); Chojnacki, 1983 a (p.435, length-weight relation); Chojnacki & Hussein, 1983 (p.53, Table 3, length-weight); d'Elbée, 1984 (p.24, Fig.3); Lam Hoi & al., 1985 (p.451, 460); Kuosa, 1989 (p.47, O2 vs. activity and survival); Jouffre & al., 1991 (p.489, lagoon); Viitasalo, 1992 (tab. 2); Castel, 1993 (p.145, long-term annual distribution); ? Hwang & Choi, 1993 (tab.3); Sellner & al., 1994 (p.249); Tanskanen, 1994 (p.397, carbon seasonal variability); Irigoien & Castel, 1995 (p.115, productivity); Irigoien & al., 1996 (p.163, gut clearance rate); Madhupratap & al., 1996 (p.77, Table 2: resting eggs); Marcus, 1996 (p.143); Meyer-Harms & von Bodungen, 1997 (p.181, feeding); Mauchline, 1998 (tab.18, 35, 40, 47, 61); Uriarte & al., 1998 (p.197, egg production, population development); Koski & Kuosa, 1999 (p.1779, egg production); Pinchuk & Paul, 2000 (p.4, table 1, % occurrence); Belmonte & Potenza, 2001 (p.173); Vieira & al., 2003 b (p.199); Lo & al., 2004 (p.89, tab.1); Katajisto, 2004 (p.751, dormancy and survival subitaneous eggs vs. anoxia & hypoxia); Hsiao & al., 2004 (p.325, tab.1); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Lan & al., 2004 (p.332, tab.1, tab.2); Uriarte & Villate, 2005 (p.863, tab.I); David & al., 2005 (p.171); Uriarte & Villate, 2006 (p.565, resting eggs); Hansen F.C. & al., 2006 (p.39, production); Schulz J. & al., 2007 (p.47, vertical zonation analysis); Chojnacki & al., 2007 (p.42, Table 2); David & al., 2007 a (p.429, figs.2, 4, 5, Table 3, 4, 5, 6); Youn & Choi, 2007 (p.222: Table1, egg production); Holmborn & Gorokhova, 2008 (p.483); Lan Y.C. & al., 2008 (p.61, Table 1, % vs stations); Li J. & al., 2008 (p.95, effects of different diets); Holeton & al., 2009 (p.661, astaxanthin storage); Holmborn, 2009 (p.12); Telesh & al., 2009 (p.18: Table 2.1); Brylinski, 2009 (p.253: Tab.1, p.256: Rem.); Martynova & al., 2009 (p.133, C/N contents, length, starvation experiments); Sun & al., 2010 (p.1006, Table 2: Yellow Sea); Mazzocchi & Di Capua, 2010 (p.423); Dvoretsky & Dvoretsky, 2010 (p.991, Table 2); Hsiao S.H. & al., 2011 (p.475, Appendix I); Sun X.-H. & al., 2011 (p.741, figs. 5.3, 4, tab.I, II, production); Martynova & al., 2011 (p.1175, seasonal abundance); Dvoretsky & Dvoretsky, 2011 b (p.469, Table A, abundance, biomass); Vehmaa & al., 2011 (p.134, algae diets vs egg production); Sopanen & al., 2011 (p.1564, mortality vs algal toxicity); Postel, 2012 (p.327, Table 1, Fig.6); Schulz J. & al., 2012 (p.3, abundance vs. hydrography); Vehmaa & al., 2012 (p.1, Table 1, egg production, egg hatching vs pH); Huo & al., 2012 (p.1, Table 1: dominance); Li C. & al., 2013 (p.101, feeding); Ben Ltaief & al., 2017 (p.1, Table III, Summer relative abundance); Baumgartner & Tarrant, 2017 (p.387, resting eggs); Belmonte, 2018 (p.273, Table I: Italian zones)Richirt & al., 2019 (p.3, Table 1, fig.2, 3, 4, 5, abundance changes vs years 1998-2014, table 2: diversity index )


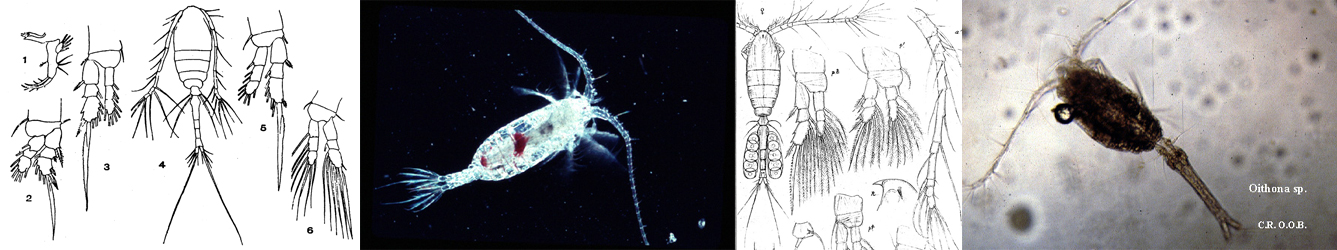

 bifilosa - Plate 1 of morphological figures',750,500);)
 bifilosa - Plate 2 of morphological figures',750,500);)
 bifilosa - Plate 3 of morphological figures',750,500);)
 bifilosa - Plate 4 of morphological figures',750,500);)
 bifilosa - Plate 5 of morphological figures',750,500);)
 bifilosa - Plate 6 of morphological figures',750,500);)
 bifilosa - Plate 8 of morphological figures',750,500);)
 bifilosa - Plate 9 of morphological figures',750,500);)
 bifilosa - Plate 10 of morphological figures',750,500);)
 bifilosa - Plate 12 of morphological figures',750,500);)
 bifilosa - Plate 13 of morphological figures',750,500);)
 bifilosa - Plate 14 of morphological figures',750,500);)


 bifilosa - Distribution map 4',650,500);)
 bifilosa - Distribution map 5',750,500);)
 bifilosa - Distribution map 6',750,500);)
 bifilosa - Distribution map 7',750,500);)
 bifilosa - Distribution map 8',750,500);)
 bifilosa - Distribution map 9',750,500);)
 bifilosa - Distribution map 11',750,500);)
 bifilosa - Distribution map 14',750,500);)
 bifilosa - Plate 1 of morphological figures){kind=link}
 bifilosa - Plate 2 of morphological figures){kind=link}
 bifilosa - Plate 3 of morphological figures){kind=link}
 bifilosa - Plate 4 of morphological figures){kind=link}
 bifilosa - Plate 5 of morphological figures){kind=link}
 bifilosa - Plate 6 of morphological figures){kind=link}
 bifilosa - Plate 7 of morphological figures){kind=link}
 bifilosa - Plate 8 of morphological figures){kind=link}
 bifilosa - Plate 9 of morphological figures){kind=link}
 bifilosa - Plate 10 of morphological figures){kind=link}
 bifilosa - Plate 11 of morphological figures){kind=link}
 bifilosa - Plate 12 of morphological figures){kind=link}
 bifilosa - Plate 13 of morphological figures){kind=link}
 bifilosa - Plate 14 of morphological figures){kind=link}
 bifilosa - Distribution map 4){kind=link}
 bifilosa - Distribution map 5){kind=link}
 bifilosa - Distribution map 6){kind=link}
 bifilosa - Distribution map 7){kind=link}
 bifilosa - Distribution map 8){kind=link}
 bifilosa - Distribution map 9){kind=link}
 bifilosa - Distribution map 10){kind=link}
 bifilosa - Distribution map 11){kind=link}
 bifilosa - Distribution map 12){kind=link}
 bifilosa - Distribution map 13){kind=link}
 bifilosa - Distribution map 14){kind=link}