|
|
 |
|
Calanoida ( Order ) |
|
|
|
Arietelloidea ( Superfamily ) |
|
|
|
Augaptilidae ( Family ) |
|
|
|
Haloptilus ( Genus ) |
|
|
| |
Haloptilus oxycephalus (Giesbrecht, 1889) (F,M) | |
| | | | | | | Syn.: | Hemicalanus oxycephalus Giesbrecht,1889; 1892 (p.384, 398, 773, Descr.F, figs.F);
? Haloptilus acutifrons : Grice, 1962 (p.223, figs.F, Rem.) | | | | Ref.: | | | Giesbrecht & Schmeil, 1898 (p.119, Rem. F); Wolfenden, 1911 (p.324); Sars, 1925 (p.252, figs.F); Farran, 1929 (p.209, 268); Rose, 1933 a (p.214, figs.F); Mori, 1937 (1964) (p.77, figs.F); Sewell, 1947 (p.194); Vervoort, 1951 (p.142, Rem.); 1957 (p.136, Descr.M, figs.M); Chiba & al., 1957 a (p.11); Brodsky, 1962 c (p.132, figs.F); Tanaka, 1964 b (p.43); Vervoort, 1965 (p.129, Rem.); Owre & Foyo, 1967 (p.82, figs.F); Ramirez, 1989 (p.81, figs.F, Rem.); Bradford, 1970 (p.347); 1970 a (p.359, figs.F, Rem.); 1971 b (p.27, figs.F,M, Rem.); Minoda, 1971 (p.40); Tanaka & Omori, 1971 (p.254); Kos, 1972 (Vol. I, figs.F, Rem.); Bradford, 1972 (p.46, figs.F); Matthews, 1972 (p.52); Björnberg, 1981 (p.654, figs.F); Gardner & Szabo, 1982 (p.388, figs.F, M); Chahsavar-Archad & Razouls, 1982 (p.38); Park, 1988 (p.9, Descr. F,M, figs.F,M, Rem.); Huys & Boxshall, 1991 (p.78, fig.F); He & al., 1992 (p.250); Razouls, 1994 (p.172, figs.F,M); Chihara & Murano, 1997 (p.728, Pl.61,62: F,M); Bradford-Grieve & al., 1999 (p.883, 941, figs.F,M); Bradford-Grieve,1999 b (p.63, figs.F,M, Rem., figs.173, 190); Conway & al., 2003 (p.86, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.246, figs.F,M, Rem.) | 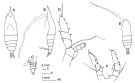 issued from : J.M. Bradford-Grieve in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). NIWA Biodiversity Memoir, 111, 1999. [p.64, Fig.35]. Female: A, habitus (dorsal); B, idem (left lateral side); C, urosome (left lateral side); D, P5. Male: E, habitus (dorsal); F, P5 (endopds: incomplete).
|
 issued from : T. Park in Antarct. Res. Ser. Washington, 1988, 47. [p.11, Fig.5]. Female: a, habitus (lateral left side); b, idem (dorsal); c, forehead (ventral); d, idem (lateral right side); e, last thoracic segment and urosome (dorsal); f, idem (lateral right side); g, Md (biting edge); h, Mx1; i, Mx2; j, P5 (posterior). Nota: Mx2: 1st to 6th protopodal lobes with 3+2+2+2+2+4 setae (instead 3 on the 6th in H. ocellatus.
|
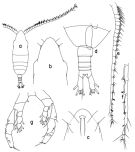 issued from : T. Park in Antarct. Res. Ser. Washington, 1988, 47. [p.13, Fig.6]. Male: a, habitus (dorsal); b, forehead (dorsal); c, idem (ventral); d, last thoracic segment and urosome (dorsal); e, left A1; f, distal part of left A1; g, P5 (posterior).
|
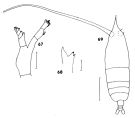 issued from : J.M. Bradford in N.Z. Jl Mar. Freshw. Res., 1970, 4 (4). [p.360, Figs 67-69]. Female (off Kaikoura, New Zealand): 67, Md (mandibular palp); 68, Md (cutting edge); 69, habitus (dorsal). Scale bars represent 1.0 mm in 69 and 0.1 mm in 67-68. Nota: Compared to H. acutifrons thie species has a longer, anteriorly pointed head, shorter A1, and oviducts which exted further, as far as the A1.
|
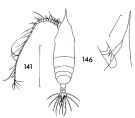 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.86, Lam. XVII, figs.141, 146 ]. Female (from off Mar del Plata): 141, habitus (dorsal); 146, forehead (lateral) Scale bars in mm: 2 (141); 0.04 (146).
|
 Issued from : G.O. Sars in Résult. Camp. Scient. Prince Albert I, 69, pls.1-127 (1924). [Pl.LXXIV, figs.12-16]. Female: 12, habitus (dorsal); 13, forehead (lateral); 14, Md; 15, Mx1; 16, Mxp.
|
 issued from : W. Vervoort in B.A.N.Z. Antarctic Reseach Expedition, Report Ser. B, Vol. III. Copepods. 1957 [Fig.128] Male (from 64°55'S, 54°35'E): a, frontal part of cephalothorax (lateral); b-c, posterior part cephalothorax and urosome (lateral and dorsal, respectively); d, forehead (dorsal); e, right Md (mandibular blade); f, Md (bitting edge) ; g-i, laft A1.
|
 issued from : W. Vervoort in B.A.N.Z. Antarctic Reseach Expedition, Report Ser. B, Vol. III. Copepods. 1957 [Fig.129] Male: a, left A2; b, right Mx1); c, right Mx2; d, right Mxp.
|
 issued from : W. Vervoort in B.A.N.Z. Antarctic Reseach Expedition, Report Ser. B, Vol. III. Copepods. 1957 [Fig.130]. Male: a, left P1 (posterior); b, right P2 (posterior); c, left P3 (posterior); d, right P4 (posterior); e, P5 (lt = left leg; rt = right leg).
|
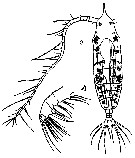 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.40, Figs.1-2]. Female: 1, Mx2; 2, habitus (dorsal).
|
 issued from : J.M. Bradford in Mem. N. Z. Oceonogr. Inst., 1972, 54. [p.49, Fig.13, (1-2]. Female (from Kaikoura, New Zealand): 1, habitus (dorsal); 2, Mxp. Scale bars1, mm (1); 0.1 mm (2).
|
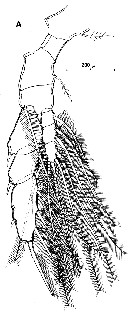 issued from : R. Huys & G.A. Boxshall in Copepod Evolution. The Ray Society, 1991, 159. [p.78, Fig.2.2.25, A]. Female from Terra Nova Expedition, 1910): P5.
|
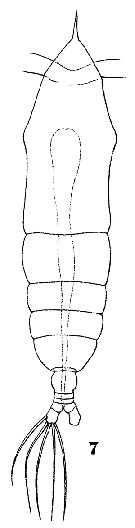 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.42, Fig.7]. As Hemicalanus oxycephalus. Female: 7, habitus (dorsal).
|
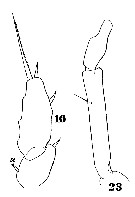 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.42, Figs.16, 23]. As Hemicalanus oxycephalus. Female: 16, exopodal segments 2 and 3 of P5; 23, endopod of A2. Si = inner seta.
|
 issued from : J.M. Bradford-Grieve, E.L. Markhaseva & C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, Edit. D. Boltovskoy. 1999. Vol.2. Copepoda. [p.1044, Fig. 7.277]. Female: Mx1. Nota: Exopodite of Mx1 with 9 setae; outer lobe 1 with 8 setae.
|
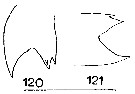 Issued from : J.M. Bradford in N.Z. Oceanogr. Inst., 1971, 206, Part 8, No 59. [p.27, Figs.120, 121]. Female (from Ross Sea): 120, right Md (edge of gnathobase); 121, left Md (edge of gnathobase). Scale bar: 100 µm.
|
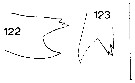 Issued from : J.M. Bradford in N.Z. Oceanogr. Inst., 1971, 206, Part 8, No 59. [p.27, Figs.122, 123]. Male (from Ross Sea): 122, left Md (ege of gnathobase); 123, right Md (edge of gnathobase). Scale bar: 100 µm.
|
 Issued from : J.M. Bradford in N.Z. Oceanogr. Inst., 1971, 206, Part 8, No 59. [p.27, Fig.124]. Male (from Ross Sea): 124, P5.. Scale bar: 100 µm. Nota: P5 is as Vervoort (1957) figures it, but Bradford includes details of ''glandular pore'' position which is at the base of every exopod spine.
|
 Issued from : C. Razouls in Ann. Inst. océanogr., Paris, 1994, 70 (1). [p.172]. Caractéristiques morphologiques de Haloptilus oxycephalus femelle et mâle adultes. Terminologie et abbréviations: voir à Calanus propinquus.
|
 Issued from : M.C. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. After Brodsky, 1962. Female: 1, habitus (dorsal); 2, forehead and rostrum (lateral); 3, A1; 4, abdomen (ventral); 5, gnathobase of Md; 6, P5.
| | | | | Compl. Ref.: | | | Hardy & Gunther, 1935 (1936) (p.182, Rem.); Wilson, 1942 a (p.189); Lysholm & al., 1945 (p.37); Sewell, 1948 (p.329, 504, 509, 514, 522, 526, 530, 532, 548, 558, 567, 568); C.B. Wilson, 1950 (p.237); Baker, 1954 (p.203, 211, fig.5); Yamazi, 1958 (p.151, Rem.); V.N. Greze, 1963 a (tabl.2); De Decker & Mombeck, 1964 (p.12); Shmeleva, 1964 a (p.1068); Furuhashi, 1966 a (p.295, vertical distribution in Oyashio region, Table 6, 7); Mazza, 1966 (p.72); De Decker, 1968 (p.45); Morris, 1970 (p.2301); Björnberg, 1973 (p.347, 386); Deevey & Brooks, 1977 (p.156, tab.2, Station "S"); Carter, 1977 (1978) (p.36); Kovalev & Shmeleva, 1982 (p.84); Lozano Soldevilla & al., 1988 (p.60); Deevey, 1971 (p.224); Dessier, 1979 (p.206); De Decker, 1984 (p.317); Cummings, 1984 (p.163, Table 2); Guangshan & Honglin, 1984 (p.118, tab.); Ali-Khan & Ali-Khan, 1984 (p.309); Longhurst, 1985 (tab.2); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Madhupratap & Haridas, 1986 (p.105, tab.1); Zmijewska, 1987 (tab.2a); Hopkins & Torres, 1988 (tab.1); Ward, 1989 (tab.2); Atkinson & al., 1990 (p.1213, tab.1); Heinrich, 1990 (p.18); Suarez & Gasca, 1991 (tab.2); Suarez, 1992 (App.1); Hosie & Cochran, 1994 (tab.2); Shih & Young,1995 (p.68); Errhif & al., 1997 (p.422); Hure & Krsinic, 1998 (p.102); Gilabert & Moreno, 1998 (tab.1, 2); Suarez-Morales & Gasca, 1998 a (p108); Siokou-Frangou, 1999 (p.478); Voronina & Kolosova, 1999 (p.71); Ansorge & al., 1999 (p.135, Rem.: p.143); Atkinson & Sinclair, 2000 (p.46, 50, 51, 54, 55, zonal distribution); Razouls & al., 2000 (p.343, tab. 5, Appendix); Pakhomov & al., 2000 (p.1663, Table 2, transect Cape Town-SANAE antarctic base); Lopez-Salgado al., 2000 (tab.1); Chiba & al., 2001 (p.95, tab.4, 7); Hsiao & al., 2004 (p.325, tab.1); Hunt, 2004 (p.1, 47, Table 3.2); Shimode & al., 2005 (p.113 + poster); Tsujimoto & al., 2006 (p.140, Table1); Dur & al., 2007 (p.197, Table IV); Schnack-Schiel & al., 2008 (p.1045: Tab.2); Morales-Ramirez & Suarez-Morales, 2008 (p.518); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Lan Y.C. & al., 2008 (p.61, Table 1, % vs stations); Galbraith, 2009 (pers. comm.); Park & Ferrari, 2009 (p.143, Table 4, 7: common deep water species, fig.1, Appendix 1, biogeography); C.E. Morales & al., 2010 (p.158, Table 1); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Hidalgo & al., 2010 (p.2089, Table 2); Mazzocchi & Di Capua, 2010 (p.424); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Swadling & al., 2010 (p.887, Table 2, 3, A1, fig.6, abundance, indicator species); Hsiao S.H. & al., 2011 (p.475, Appendix I); Swadling & al., 2011 (p.118, Table 2, interannual abundance); Michels & al., 2012 (p.369, Table 1, occurrence frequency); Belmonte & al., 2013 (p.222, Table 2, abundance vs stations); Ojima & al., 2013 (p.1293, Table 2, 3, abundance); Lee D.B. & al., 2013 (p.1215, Table 1, abundance, composition); Lidvanov & al., 2013 (p.290, Table 2, % composition); Zaafa & al., 2014 (p.67, Table I, occurrence); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); El Arraj & al., 2017 (p.272, table 2, seasonal composition); Belmonte, 2018 (p.273, Table I: Italian zones); Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions). | | | | NZ: | 19 | | |
|
Distribution map of Haloptilus oxycephalus by geographical zones
|
| | | | | | | | | | | | | | | 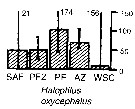 Issued from : A. Atkinson & J.D. Sinclair in Polar Biol., 2000, 23. [p.50, Fig.3] Issued from : A. Atkinson & J.D. Sinclair in Polar Biol., 2000, 23. [p.50, Fig.3]
Haloptilus oxycephalus from Scotia Sea.
Median and interquartile ranges of copepods (nos /m2) in the five water zones; from north to south these are SAF Subantractic Front area, PFZ Polar frontal Zone, PF Polar Front area, AZ Antarctic Zone, WSC Weddell-Scotia Confluence area/ East Wind Drift.
Numbers on the plots are upper interquartiles where these could not be scaled. |
 issued from A. de C. Baker in 'Discovery' Rep., 1954, 27. [p.215, Fig.5]. issued from A. de C. Baker in 'Discovery' Rep., 1954, 27. [p.215, Fig.5].
Occurrence of Haloptilus oxycephalus in all longitudes around Antarctic zone of the Southern Ocean.
The percentage frequency of occurrence in samples taken within every 20° of longitude.
Nota: During the 'Discovery' investigations some thousands of plankton samples have been taken from stations spread over the whole of the Southern Ocean at all seasons of the year, the majority south of the Antarctic Convergence. An arbitrary selection of samples has been made from hauls between the surface and a depth of 250 m, which means that they have been taken from within the limits of the Antarctic surface water.
|
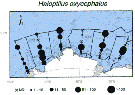 Issued from : K.M. Swadling, So. Kawaguchi & G.W. Hosie in Deep-Sea Research II, 2010, 57. [p.897, Fig.6 (continued)]. Issued from : K.M. Swadling, So. Kawaguchi & G.W. Hosie in Deep-Sea Research II, 2010, 57. [p.897, Fig.6 (continued)].
Distribution of indicator species Haloptilus oxycephalus from the BROKE-West survey (southwest Indian Ocean) during January-February 2006.
Sampling with a RMT1 net (mesh aperture: 315 µm), oblque tow from the surface to 200 m.
The survey area was located predominantly within the seasonal ice zone, and in the month prior to the survey there was considerable ice coverage over the western section but none over the east.
See map showing sampling sites in Calanus propinquus. |
| | | | Loc: | | | Antarct. (Amundsen Sea, Peninsula, SW & SE Atlant., South Georgia, Weddell Sea, Indian, Dumont d'Urville Sea, Lützow-Holm Bay, Sw & SE Pacif.), South Georgia, sub-Antarct. (SW Atlant., Indian, SW & SE Pacif.), South Africa (E & W), Patagonia, Congo, SE Atlantic, G. of Guinea, S Cape Verde Is., S Brazil, off Amazon, Caribbean Colombia, Caribbean, G. of Mexico, Cuba, Florida, Sargasso Sea, off Bermuda (Station "S"), off S Iceland, Morocco-Mauritabia, Canary Is., off W Tangier, Medit. (Algiers Bay, Baleares, Ligurian Sea, Tyrrhenian Sea, Taranto, Adriatic Sea, Ionian Sea, Saronikos Gulf), Arabian Sea, Pakistan, Natal, Indian, off SE Madagascar, China Seas (East China Sea, South China Sea), Taiwan (SW, E), Japan, Tanabe Bay, Bering Sea, S Aleutian Is., Vancouver Is., Bikini, Hawaii, Pacif. (tropical), Pacif. (W equatorial), New Zealand (Kaikoura, SE South Island, SE North Island), Tasman Sea, off W Guatemala, W Costa Rica, off W Galapagos, SE Pacif. (tropical), Peru, Chilie (N-S, Concepcion) | | | | N: | 114 | | | | Lg.: | | | (1) F: 3,5; (16) F: 3,7-3,3; M: 2,5; (25) F: 4,95-3,83; M: 2,57; (31) F: 4,9-4,7; (35) F: 4,08-4,02; (46) F: 3,8; (69) F: 3,56; (90) F: 5-3,64 ; ? (101) F: 3,8; (102) F: 3,1; M: 2,85; (116) F: 3,64; (150) F: 3,25; (208) F: 5,2-4,57; (254) F: 4,25; (313) F: 4,4; 4,3; (866) F: 3-5; M: 2,44-2,85; (909) F: 3,3-4,1; M: 2,4-2,6; (991) F: 3-4,1; M: 2,4-2,85; (1230) F: 3,0-3,5; {F: 3,00-5,20; M: 2,40-2,85} | | | | Rem.: | epi-bathypelagic. Reported at 100 m depth or less but also recorded down to 600 m (Matthews, 1972). Sampling depth (Antarct., sub-Antarct.): 0-1000 m. Sargasso Sea: 0-1500 m (Deevey & Brooks, 1977, Station "S");
According to Vervoort (1965) the locality records in the NW Pacific could correspond to Haloptilus pseudooxycephalus. See remarks in Bradford-Grieve (1999 b, p.60).
For Park (1988, p.14) this species in habitus and anatomical details of the appendages of both the female and the male is very close to H. ocellatus, but can be distinguished from it by the presence of 4 setae on the 6th protopodal lobe of Mx2.
For Mazzocchi & al. (1995, p.31), the species resembles H. acutifrons, the latter of which is rarer from Antarctic and sub-Antarctic seas. As discussed by Park (1988), the two species are practically indistinguishable; the only useful anatomical detail to determine acutifrons is the relatively shorter cephalic spine.
See in DVP Conway & al., 2003 (version 1) | | | Last update : 24/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed March 24, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
