|
|
 |
|
Calanoida ( Order ) |
|
|
|
Calanoidea ( Superfamily ) |
|
|
|
Calanidae ( Family ) |
|
|
|
Cosmocalanus ( Genus ) |
|
|
| |
Cosmocalanus darwini (Lubbock, 1860) (F,M) | |
| | | | | | | Syn.: | Undina darwini Lubbock,1860; Brady, 1883 (p.54, figs.F,M);
Calanus darwinii : Giesbrecht, 1892 (p.91, 127, figs.F,M); Giesbrecht & Schmeil, 1898 (p.17); Thompson & Scott, 1903 (p.232, 241); Cleve, 1904 a (p.185); Pesta, 1913 (p.30, fig.M); Farran,1929 (p.207, 217); Dakin & Colefax, 1933 (p.203); Mori, 1937 (1964) (p.18, figs.F,M); Chiba, 1953 (p.235, Rem.: anomalies); 1957 (p.306); 1957 a (p.11); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Maiphae & Sa-ardrit, 2011 (p.641, Table 2);
Calanus darwini : Carl, 1907 (p.16);
Undinula darwini : A. Scott, 1909 (p.17, Rem.); Sewell, 1912 (p.353, 356); 1914 a (p.197, Rem.); 1929 (p.42, Rem.: 3 forms); Farran, 1936 a (p.78); Wilson, 1936 c (p.90); Dakin & Colefax, 1940 (p.89, figs.F,M); Wilson, 1942 a (p.210); Vervoort, 1946 (p.77, Rem.); Sewell, 1947 (p.18, Rem.F); 1948 (p.322, 357, 390, 395, 403, 413, 414, 417, 422, 433, 436, 439, 442, 453, 459, ); 1951 (p.366, figs.F, Rem.: parasites); C.B. Wilson, 1950 (p.349, figs.F,M); Krishnaswamy, 1953 (p.110); Rose, 1956 (p.459); Tanaka, 1956 (p.265, Rem.); Kott, 1957 (p.5, Rem.: p.8); Yamazi, 1958 (p.147, Rem.); Tanaka, 1960 (p.20); Heinrich, 1961 b (p.93); Fagetti, 1962 (p.8); Ganapati & Shanthakumari, 1962 (p.7, 16); Grice, 1962 (p.178, figs.F, Rem.); Kasturirangan, 1963 (p.14, figs.F,M); Brodsky, 1962 c (p.102, figs.F,M); De Decker, 1964 (p.16, 25, 27); De Decker & Mombeck, 1964 (p.14); Chen & Zhang, 1965 (p.31, 129, figs.F, M); Saraswathy, 1966 (1967) (p.75); Fleminger, 1967 a (tabl.1); Grice & Hulsemann, 1967 (p.13); De Decker, 1968 (p.45); Vidal, 1968 (p.17, figs.F,M); Vinogradov, 1968 (1970) (p.78, 79, 80, 83, 278); Itoh, 1970 (tab.1); Timonin, 1971 (p.281, trophic group); Shih & al., 1971 (p.203); Brodsky, 1972 (p.256); Brodsky, 1972 (1975) (p.73,113, 116, 117, figs.F,M); Grice, 1972 (p.178, figs.F, Rem.); Heinrich, 1973 (p.95, fig.2); Björnberg, 1973 (p.283, 389); Marques, 1976 (p.986); Timonin, 1976 (p.79, vertical distribution); Tranter, 1977 (p.596); Timonin & Voronina, 1977 (p.283); Carter, 1977 (1978) (p.35); Frontier, 1977 a (p.14); Hayward, 1980 (p.295, Table 2, vertical distribution, feeding); Sreekumaran Nair & al., 1981 (p.493), Björnberg & al., 1981 (p.618, figs.F,M); Gaudy & Boucher, 1983 (p.37, Table 1, 2, 3, 4, 5, fig.1, 2, Rem.: metabolism); Dessier, 1983 (p.89, Tableau 1, 2, Rem. p.98, 99, %); Binet,1984 (tab.3); 1985 (p.85, tab.3); Guangshan & Honglin, 1984 (p.118, tab.); De Decker, 1984 (p.315, 366: chart); Sazhina, 1985 a (p.491, tab.1); Petipa & Borichenko, 1985 (tab.2); Sarkar & al., 1986 (p.178); Brinton & al., 1986 (p.228, Table 1); Renon, 1987 (tab.2); Ambler & Miller, 1987 (tab.2, 3, 4, 5); Jimenez-Perez & Lara-Lara, 1988; Dessier, 1988 (tabl.1); Hernandez-Trujillo, 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Afanas'yev & al., 1989 (p.225, genetic); Herman, 1989 (p.247); Suarez & al., 1990 (tab.2); Hattori, 1991 (tab.1, Appendix); Yoo, 1991 (tab.1); Huntley & Lopez, 1992 (p.201, Table B1, growth rate, temperature-dependent production); He & al., 1992 (p.250); Kim & al., 1993 (p.270); Go & al., 1997 (tab.1); Hwang & al., 1998 (tab.II); Noda & al., 1998 (p.55, Table 3, occurrence); Hernandez-Trujillo, 1999 (p.284, tab.1); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); Fernandez-Alamo & al., 2000 (p.1139, Appendix); Suarez-Morales al., 2000 (p.751, tab.1); Hernandez-Trujillo & Esqueda-Escarcega, 2002 (in Appendix); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Lavaniegos & Jiménez-Pérez, 2006 (p.158, tab.2, 4, Rem.); Rakhesh & al., 2006 (p.93, Table 2, spatial distribution); McKinnon & al., 2008 (p.843, Tab.I, p.848: Tab. IV); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); W.-B. Chang & al., 2010 (p.735, Table 2, 3, fig.5, abundance); Tutasi & al., 2011 (p.791, Table 2, abundance distribution vs La Niña event); Jungbluth & Lenz, 2013 (p.630, Table I); Peijnenburg & Goetze, 2013 (p.2765, genetic data);
Cosmocalanus darwini s.l. : Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution as Cosmocalanus spp., Rem.: p.225); | | | | Ref.: | | | Bradford & Jillett, 1974 (p.6, 13, figs.F,M); Greenwood, 1976 (p.8, figs.F); Fleminger, 1985 (p.276, 284, Table 1, 4, fig.M, Rem.: A1); Bradford, 1988 (Rem., p.74, 76); Nishida, 1989 (p.173, table 2, no M dorsal hump); Hirakawa, 1991 (p.376: fig.2); Bradfod-Grieve, 1994 (p.35, figs.F,M, fig.98); Chihara Murano, 1997 (p.737, Pl.63: F,M); Barthélémy, 1999 a (p.10, Fig.14, A, E); Bradford-Grieve & al., 1999 (p.877, 906, 907, figs.F,M); Bucklin & al., 2003 (p.335, tab.2, fig. 4, Biomol.); Conway & al., 2003 (p.145, figs.F,M, Rem.); Boxshall & Halsey, 2004 (p.81: figs.F,M); Mulyadi, 2004 (p.8, 20, figs.F,M, Rem.); Phukham, 2008 (p.112, figs.M); Lacuna & al., 2013 (p.74, Figs.F,M, Rem.) | 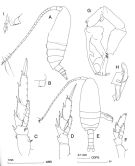 issued from : J.M. Bradford-Grieve in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). New Zealand Oceanographic Institute Memoir, 102, 1994. [p.37, Fig.13]. Female: A, habitus (left lateral side); B, genital somite and pedigerous segment 5 (right side); C, P3; D, P5; I, genital somite and pedigerous segment 5 from a another specimen (different station). Nota: Inner edge of coxa of P5 with teeth. Basis of P2-P3 with posterior surface spinules. Male: E, habitus (dorsal); F, right P5: G, left P5; H, detail of the distal part of left P5.
|
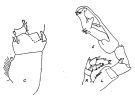 issued from : J.M. Bradford & J.B. Jillett in Crustaceana, 1974, 27 (1). [p.14, Fig.2, C,J]. Female: C, P2. Male: J, P5.
|
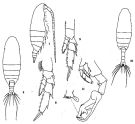 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.4, 6-11]. As Undinula darwinii. Female (from E China Sea): 6, habitus (dorsal); 7, idem (lateral right side); 8, P3 (posterior); 9, P5 (posterior). Male: 10, habitus (dorsal); 11, P5 (posterior).
|
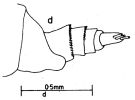 issued from : J.G. Greenwood in Proc. R. Soc. Qd, 1976, 87. [p.19, Fig.3, d]. Female (from Moreton Bay, E Australia): d, posterior metasomal segment and urosome (lateral left side).
|
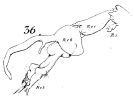 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.8, Fig.37]. Male: 37, P5.
|
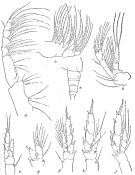 issued from : R.B.S. Sewell in Mem. Indian Mus., 1929, X. [p.44, Fig.11]. As Undinula darwini forma typica. Female (from N Indian Ocean): a, posterior thoracic margin and abdomen (lateral right side); b, A2; c, Mx2; d, Mxp; e, P1; f, P2; g, P2 abnormal (endopod of 2 segments only); h, P3; j, P5.
|
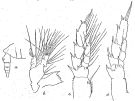 issued from : R.B.S. Sewell in Mem. Indian Mus., 1929, X. [p.46, Fig.12]. Undinula darwini var. intermedia. Female (from N Indian Ocean): a, posterior thoracic margin and urosome (lateral left side); b, Md; c, P3; d, P5.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.5, Figs.6-12]. As Calanus darwinii. Female: 6-7, habitus (lateral and dorsal, respectively); 8, P5; 11, P3; 12, P2. Male: 9, P5; 10, habitus (lateral).
|
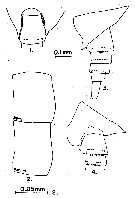
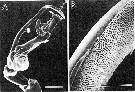 issued from : S. Ohtsuka & R. Huys in Hydrobiologia, 2001, 453/454. [p.453, Fig.8]. Male (from Japan): A, left exopodal segments (arrowed part magnified in B); B, magnification of terminal inner surface of outer process on 2nd segment. Scale bars: 0.1 mm (A); 0.01 mm (B). Nota: The 2nd exopodal segment of the left P5 bears patches of fine granules which may also function ib graping the spermatophore.
|
 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.179, Pl.2, Figs.1-4]. As Undinula darwini f. typica. Female (from equatorial Pacific): 1, posterior part of thorax and genital segment (dorsal); 2, segments 6 and 7 of A1; 3-4, posterior part of thorax and urosome ( left and right side, respectively).
|
 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.179, Pl.2, Figs.5-6]. As Undinula darwini f. symmetrica. Female (from equatorial Pacific): 5-6, posterior part of thorax and genital segment (right and left side, respectively).
|
 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.179, Pl.2, Figs.7-9]. As Undinula darwini f. intermedia. Female (from equatorial Pacific): 7-8, posterior part of thorax and urosome (left side); 9, same (right side).
|
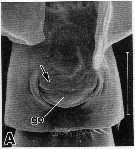 issued from : R.-M. Barthélémy in These Doct. Univ. Provence (Aix-Marseille I), 1999. [Fig.14, A]. Female (from 30°45'S, 173°16'E): A, external ventral view genital double-somite. go = genital operculum; Note the anterior thick pad (arrow). Scale bar: 0.100 mm.
|
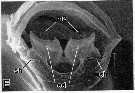 issued from : R.-M. Barthélémy in These Doct. Univ. Provence (Aix-Marseille I), 1999. [Fig.14, E]. Female: internal genital area. m2 = egg-laying ducts muscles; ed = egg-laying duct; sh = shell ducts. Scale bar: 0.020 mm.
|
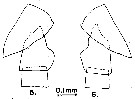
 issued from : A. Fleminger in Mar. Biol., 1985, 88. [p.284, Fig.6 B]. Male (from N Pacific, subarctic): Left A1 proximal segments (ventral view); Nota: see remarks in Calanus s.l. pacificus californicus (Fleminger, 1985, p.275) concerning the dimorphism in the female A1.
|
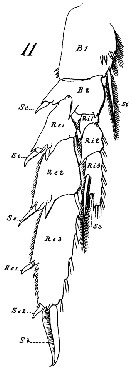
 Issued from : G.S. Brady in Rep. Scient. Results Voy. Challenger, Zool., 1883, 8 (23). [Pl.XVI, Figs1-14]. As Undina darwinii. Female: 3, A1; 4, apical segments (magnified); 6, Md; 7, Mx1; 8, Mx2; 9, Mxp; 10, exopodite and basis of P3; 14, urosome. Male: 1, habitus (lateral); 2, A1; 11, right P5; 12, left P5; 13, urosome.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 8, Fig.11]. As Calanus darwinii. Female: 11, P3 (anterior view).
|
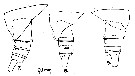
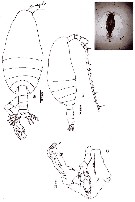 issued from : N. Phukham in Species diversity of calanoid copepods in Thai waters, Andaman Sea (Master of Science, Univ. Bangkok). 2008. [p.195, Fig.69]. Male (from W Malay Peninsula): a-b, habitus (dorsal and lateral, respectively); c, P5. Body length after the drawing: M = 1.600 mm.
|
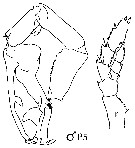 issued from : J.M. Bradford-Grieve, E.L. Markhaseva & C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, Edit. D. Boltovskoy. 1999. Vol.2. Copepoda. [p.980, Fig. 7.17]. Male: P5 (L = left leg; r = right leg). Nota: Left P5 highly modified, of complex structure (prehensile). Coxa of left and right P5 legs with teeth on the inner edge . Exopodite 2 of P2 without invaginated outer proximal border.
|
 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.21, Fig.8]. Female (from Indonesian Seas): a, habitus (dorsal); b, last thoracic segment and urosome (lateral right side). Male: c, habitus (dorsal); d, P5.
|
 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.22, Fig.9]. Female: a-e, P1 to P5, respectively: f-i, variations of left posterior ends of last thoracic segment.
|
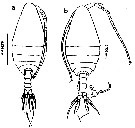 ssued from : M.L.D.G. Lacuna, D.C. Sagrado, R.O. Mejorada, D.D. Simyunn & M.J.J. Pueblos in ABAH Bioflux, 2013, 5 (1). [p.74, Fig.29]. Female (from Mindanao): a, habitus (dorsal). Male: b, habitus (dorsal). Nota: Body dark brown coloration in live specimens.
|
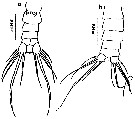 ssued from : M.L.D.G. Lacuna, D.C. Sagrado, R.O. Mejorada, D.D. Simyunn & M.J.J. Pueblos in ABAH Bioflux, 2013, 5 (1). [p.75, Fig.30]. Female : a, urosome (ventral). Male: b, urosome (dorsal). Nota: Each caudal ramus has 6 setae with the 2nd seta much longer than the rest.
|
 ssued from : M.L.D.G. Lacuna, D.C. Sagrado, R.O. Mejorada, D.D. Simyunn & M.J.J. Pueblos in ABAH Bioflux, 2013, 5 (1). [p.75, Figs.31, 32]. Female: a, left A1; b, right A1. Male: c, left A1; d, right A1. Nota: Both A1 female 25-segmented; A1 male 24-segmented
|
 ssued from : M.L.D.G. Lacuna, D.C. Sagrado, R.O. Mejorada, D.D. Simyunn & M.J.J. Pueblos in ABAH Bioflux, 2013, 5 (1). [p.76, Fig.33]. Female: a, P1; b, P2; c, P3; d, P4; e, P5.
|
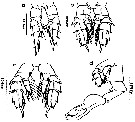 ssued from : M.L.D.G. Lacuna, D.C. Sagrado, R.O. Mejorada, D.D. Simyunn & M.J.J. Pueblos in ABAH Bioflux, 2013, 5 (1). [p.77, Fig.34]. Male: a, P2; b, P3; c, P4; d, P5. Nota: Right P5 smaller, biramous, with exopod and endopod 3-segmented. The inner margin of basipodal segment serrated. Left P5 uniramous, having 3-segmented highly modified exopod and rudimentary endopod, the inner margin of basipodal segment with spinules; the 2nd exopodal segment bears an elongated external spine with an inner-like projection, while the 3rd segment is widened and armed with a stout bifurcate process; the tooth on the inner margin of the outer arm is at or near the center.
|
 Issued from : M. Chihara & M. Murano in An Illustrated Guide to Marine Plankton in Japan, 1997. [p.742, Pl. 63, fig.14 a-e]. Female: c, P5; d, habitus (lateral); f, posterior corner of last thoracic segment and urosome (lateral). Male: e, habitus (lateral); a, P5.
|
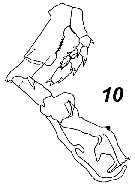 Issued from : V.N. Andronov in Russian Acad. Sci. P.P. Shirshov Inst. Oceanol. Atlantic Branch, Kaliningrad, 2014. [p.73, Fig.19, 10]. Cosmocalanus darwinii after Brodsky, 1972. Male P5.
|
 Issued from : M.C. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. As Undinula darwinii. After Brodsky, 1962. Female: 1-2, habitus (dorsal and lateral, respectively); 3-4, last thoracic segment (left side) and 1st abdominal segment ('forma b'); 5-6, same, left and right corner ('forma A'); 7, coxopodite of P5; male: 8, habitus (dorsal); 9, P5; 10, P5 (teeth on left and right basis, anterior region).
| | | | | Compl. Ref.: | | | Stephen & Iyer, 1979 (p.228, tab.1, 3, 4, figs.3, 4); Stephen, 1984 (p.161, 169, Distribution vs thermocline & geographic); Madhupratap & Haridas, 1986 (p.105, tab.1); 1990 (p.305, fig.6: vertical distribution night/day; fig.7: cluster); Heinrich, 1990 (p.16); Hirakawa & al., 1990 (tab.3); Othman & al., 1990 (p.561, 563, Table 1); Shih & Young, 1995 (p.68); Madhupratap & al., 1996 (p.866); Padmavati & Goswami, 1996 a (p.85, fig.3, Table 4, vertical distribution); Kotani & al., 1996 (tab.2); Park & Choi, 1997 (Appendix); Wong & al, 1998 (tab.2); Smith S. & al., 1998 (p.2369, Table 6, moonsoon effects); Padmavati & al., 1998 (p.349); Stephen, 1998 (p.339, 340, chart); Suarez-Morales, 1998 (p.345, Table 1); Suarez-Morales & Gasca, 1998 a (p108); Hassett & Boehlert, 1999 (p.571); Lo & al., 2001 (1139, tab.I); Madhupratap & al., 2001 (p. 1345, vertical distribution vs. O2, figs.4, 5: clusters, p.1353: Rem.); Hernandez-Trujillo & Suarez-Morales, 2002 (p.748, tab.1); Hwang & al., 2003 (p.193, tab.2); Shimode & Shirayama, 2004 (tab.2); Hsiao & al., 2004 (p.325, tab.1); Hsieh & al. 2004 (p.397,tab. 1); Lan & al., 2004 (p.332, tab.1, tab.2); Lo & al.*, 2004 (p.218, tab.1, fig.6); Gallienne & al., 2004 (p.5, tab.3); Lo & al., 2004 (p.89, tab.1); Prusova & Smith, 2005 (p.76); Zuo & al., 2006 (p.159, tab.1, 3, abundance, fig.8: stations group); Schmitt & al., 2006 (p.287, swimming behavior); Hwang & al., 2006 (p.943, tab.I); Hwang & al., 2007 (p.23); Dur & al., 2007 (p.197, Table IV); Jitlang & al., 2008 (p.65, Table 1); McKinnon & al., 2008 (p.843, fig.7: Table 1); Humphrey, 2008 (p.84: Appendix A); Morales-Ramirez & Suarez-Morales, 2008 (p.518); Fernandes, 2008 (p.465, Tabl.2); Wishner & al., 2008 (p.163, Table 2, fig.8, oxycline); Lan Y.C. & al., 2008 (p.61, Table 1, % vs stations); Tseng L.-C. & al., 2008 (p.153, Table 2, fig.5, occurrence vs geographic distribution, indicator species); Tseng & al., 2008 (p.402, Table 2); C.-Y. Lee & al., 2009 (p.151, Tab.2); Tseng & al., 2009 (p.327, fig.5, feeding); W Zhang & al., 2009 (p.266: table 2); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Cornils & al., 2010 (p.2076, Table 3); Xu & Gao, 2011 (p.514, figs.3, 4, Table 2: optimal salinity); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Pillai H.U.K. & al., 2011 (p.239, Table 3, vertical distribution); Nowaczyk & al;, 2011 (p.2159, Table 2: in Mediterranean Sea); Hsiao S.H. & al., 2011 (p.475, 481: indicator species, Appendix I); Hsiao & al., 2011 (p.232, Table 1: abundance, %, Table 2: trace metal concentration, vs transect); Hsiao & al., 2011 (p.317, Table 2, indicator of seasonal change); Guo & al., 2011 (p.567, table 2, indicator); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); DiBacco & al., 2012 (p.483, Table S1, ballast water transport): Johan & al., 2012 (2013) (p.1, Table 1); Mulyadi & Rumengan, 2012 (p.202, Rem.: p.204); Tseng & al., 2012 (p.621, Table 3: abundance); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Hsiao & Fang, 2013 (p.175, Hg bioaccumulation); Tseng & al., 2013 (p.507, seasonal abundance); Tseng & al., 2013 a (p.1, Table 3, 4, abundance); Jagadeesan & al., 2013 (p.27, Table 3, seasonal abundance); Hirai & al., 2013 (p.1, Table I, molecular marker); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Zakaria & al., 2016 (p.1, Table 1, Rem.); Palomares-Garcia & al., 2018 (p.178, Table 1: occurrence); Hirai & al., 2020 (p.1, Fig. 5: cluster analysis (OTU), spatial distribution). | | | | NZ: | 16 | | |
|
Distribution map of Cosmocalanus darwini by geographical zones
|
| | | | | | | | | | | | 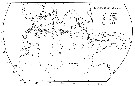 issued from : R. Stephen in Pelagic Biogeography ICoPB II. Proc. 2nd Int. Conf. Final report of SCOR/IOC working group 93. 9-14 July 1995. Workshop Rep. No.142. UNESCO, 1998. issued from : R. Stephen in Pelagic Biogeography ICoPB II. Proc. 2nd Int. Conf. Final report of SCOR/IOC working group 93. 9-14 July 1995. Workshop Rep. No.142. UNESCO, 1998.
P.340, Fig.2: Distribution of Cosmocalanus darwini in the Indian Ocean. |
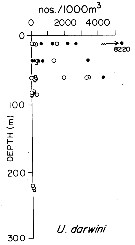 issued from : T.L. Hayward in Mar. Biol., 1980, 58. [p.299, Fig.2]. issued from : T.L. Hayward in Mar. Biol., 1980, 58. [p.299, Fig.2].
Plots of day (open circle) and night (filled circle) depth distributions of Undinula (= Cosmocalanus) darwini in the North Pacific central gyre (main sampling location: 25°N, 155°W with drogue), September 1968. |
 issued from : T.L. Hayward in Mar. Biol., 1980, 58. [p.302, Fig.4]. issued from : T.L. Hayward in Mar. Biol., 1980, 58. [p.302, Fig.4].
Day and night feeding indices (relative degree of gut fullness of copepods collected under differing environmental conditions) for females Undinula (= Cosmocalanus) darwini from N Pacific central gyre (main sampling location: 28°N, 155°W), June 1973.
Numbers in abscisse : stations. |
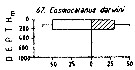 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 67- Cosmocalanus darwini.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
 Issued from : Y.-Q. Chen in CalCOFI Rep., 1986, XXVII. [Fig.8, p.216]. Issued from : Y.-Q. Chen in CalCOFI Rep., 1986, XXVII. [Fig.8, p.216].
Vertical abundance of females, males and late copepodites.
Nota: The distribution during the Krill Expedition in the Pacific east tropical (23°N to 3°S) for the two species (C. darwini and C. caroli. This category constitutes 3.65 % of the total number of copepods (samples with a bongo net (330 and 220 mesh apertures) and occurred mainly from 3°N to 4°S above 200 m at stations 20-22
The maximum number of females (17,900 individuals/1000 m3 was concentrated between 100 and 150 m. Other high concentrations above 120 m at stations 14, 12,10 between 23° and 20°N.
Although diel vertical migration was not apparent, the numbers at night were much higher than the numbers in day samples..
The species were concentrated between 50 m and the surface.
Females numbers were higher than those of males.
The mean percentage combined was 3.7% (day: 2.8, night: 4.5) for all samples .
See in Subeucalanus subtenuis figs.1, 12, 13). |
| | | | Loc: | | | South Africa (E & W), Namibia, Atlant. (tropical), S Argentina, Brazil, Caribbean, Yucatan, G. of Mexico, Bermuda, Hudson Bay (in Wilson, 1936 d), Medit. (NW, Algero- Provensal Basin, Sicily Channel, Ionian, W Egyptian coast, Lebanon Basin), Suez Canal, Red Sea, Natal, Madagascar, Nosy Bé, Rodrigues Is. - Seychelles, G. of Aden, Arabian Sea, Maldive Is., Sri Lanka, India (Goa - Gujarat, G. of Mannar, Palk Bay, Madras, Lawson's Bay, Hooghly estuary), Indian (equatorial), E India, Bay of Bengal, Nicobar Is. (Nankauri Harbour), Barren Island, S Burma, W Malay Peninsula, W Pacif. (equatorial), W Australia, Thailand, Indonesia-Malaysia, Sarawak: Bintulu coast, Java Sea, Flores Sea, Ambon Bay, N Celebes, Philippines (Mindanao), Viet-Nam (Cauda Bay), Gulf of Tonkin, China Seas (Yellow Sea, East China Sea, South China Sea), Taiwan Strait, Taiwan (S, E, SW, W, NW, N, Mienhua Canyon, NE), Kueishan Is., Okinawa, S Korea, Korea Strait, Japan Sea, Japan, off Japan (Kuchinoerabu Is., SE & NE), Bering Sea, off Kamchatka, Caroline-Marshall Is., Hawaii, Kaneohe Bay, off NE Hawaii, California, W Baja California, Gulf of California, La Paz, G. of Tehuantepec, W Mexico, W Costa Rica, Costa Rica (Dome), Clipperton Is., N Pacific (central gyre), Pacif. (equatorial), Australia (G. of Carpentaria, Great Barrier, Moreton Bay, off Sydney), New Zealand, New Caledonia, Galapagos- Zcuador, off Peru, Chile | | | | N: | 181 | | | | Lg.: | | | (28) M: 1,85-1,63; (34) F: 2,2-2,16; (35) F: 2,5-2,1; M: 2,1-1,9; (47) F: 2,2-1,6; M: 1,8-1,7; (66) F: 2,26-2; M: 2,22-1,95; (101) F: 2,07-1,8; M: 1,8-1,6; (104) F: 2,25; M: 1,8; (125) F: 2,56-2,13; M: 2,1-1,93; (150) F: 2,35-1,97; M: 1,85-1,77; (187) F: 2,58-2,45; M: 2,23-2,03; (196) F: 2,4-2,3; M: 2,35-2,03; (290) F: 2,3-1,95; M: 2,15-1,8; (333) M: 1,72; (334) F: 2,3-1,66; M: 2-1,8; (530) F: 2; M: 1,8; (786) F: 2,39-2,08; M: 2,3-1,97; (795) F: 2,5; M: 2; (864) F: 1,68-2,26; (991) F: 1,6-2,4; M: 1,63-2,05; (1122)* F: 1,25; M: 1,2; (1130) F: 1,7; M: 1,6; (1230) F: 1,8-2,4; M: 1,8-1,9; {F: 1,25-2,58; M: 1,20-2,35}
*: The lengths given by Mulyadi (2004) are exceptionally small. | | | | Rem.: | Epipelagic, sometimes deeper.
Sewell (1914 a, p.197) points to: According to Giesbrecht (1893, Pl.VII, fig.29), both 1st and 2nd abdominal segments are furnished with a row of sharp spines on the posterior margin, and the posterior thoracic margin on the left side is produced backwards in a rectangular projection; but numerous examples from various localities in the Indian Ocean has shown that these characters are subject to very considerable variation. The spinulation of the abdominal segments, many specimens possessed spines only on one of the segments, while in some case spines were absent altogether. In numerous examples the posterior thoracic margin formed almost a uniform curve, with a very slight rounded projection towards the ventral side; in other cases this prominence was more marked and formed a somewhat triangular prominence, while in a few cases the typical projection was absent. For Sewell (p.198-200) the species U. caroli is only a variety in which appears that the sole appreciable difference between these forms lies in the structure of P5 male.
Cosmocalanus darwini intermedia (Sewell,1929) (F)
Syn.: Undinula darwini intermedia Sewell, 1929 (p.45, figs.F); Vervoort, 1946 (p.80, Rem.); Sewell, 1951 (p.367); Grice, 1962 (p.180, figs.F); Kasturirangan, 1963 (p.14, fig.F); Silas, 1972 (p.644).
Ref.: Bradford-Grieve, 1994 (p.35, Rem.)
Loc.: Inde, Indo-Malaisie, Nouvelle-Zélande
Lg.: (28) F: 2,15-1,75; (101) F: 2,07-1,84
Cosmocalanus darwini typica (Sewell,1929) (F)
Syn.: Undinula darwini typica Sewell, 1929 (p.43, figs.F); Vervoort, 1946 (p.78, Rem.); Sewell, 1951 (p.367); Grice, 1962 (p.180, figs.F); Kasturirangan, 1963 (p.14, fig.F); Chen & Zhang, 1965 (p.129: Rem.)
Ref.: Bradford-Grieve, 1994 (p.35)
Loc.: Inde, Indo-Malaisie, E China Sea, Nouvelle-Zélande
Lg.: (28) F: 2,05-1,8; (29) F: 1,66; (101) F: 2,18-1,8
Cosmocalanus darwini symmetrica (Sewell,1929) (F,M)
Syn.: Undinula darwini symmetrica Sewell, 1929 (p.45); Vervoort, 1946 (p.78, Rem.); Sewell, 1951 (p.367); Grice, 1962 (p.180, figs.F); Kasturirangan, 1963 (p.14, fig.F); Vidal,1968 (p.17, figs.F,M); Silas, 1972 (p.644).
Loc.: Inde, Indo-Malaisie
Lg.: (28) F: 2,05-1,7; (101) F: 2,04-1,8; (187) F: 2,58-2,45; M: 2,23-2,03.
Nota: The prolongation of the posterior margin of the last thoracic segment on the left side is entirely absent.
Sewell (1929) reported the 3 subspecies females in rthe Indian Ocean and Grice (1962) in the equatorial Pacific. According to Mulyadi (2004) all of them were recorded only in the eastern Indonesian Seas (Wallacea region).
Sewell (1947, p.18-19) points to an abnormality: the two A1 approximately equal length, that on the right side measuring 2.05 and 2.08 mm, and that on the left 2.00 and 2.10 mm; whereas the right was normal and consisted of full 25 segments (segments 8 and 9 fused), on the left side in each specimen only 18 segments (8 and 9 fused).
Nowaczyk & al. (2011 (p.2173) reported this species for the first time in the Mediterranean Sea, both in the western and eastern basins. For the authors it is difficult to conclude about its origin in the mediterranean Sea. It is expected to undergo lesseptian introduction, but this species was found in lower abundance in the eastern basin than in the western basin.
Timonin (1971, p.282) considers the trophic interrelations in the equatorial and tropical Indian Ocean, and divides the plankters into 6 trophic groups from the litterature and the results of studies of mouth-parts structure and intestine content. This species is a fine-filter feeder.
For Itoh (1970 a, fig.2, from co-ordonates) the Itoh's index value from mandibular gnathobase = 340, and for Gaudy & Boucher (1983): 340.
See in DVP Conway & al., 2003 (version 1)
R. Stephen: Data sheets of NIO, Kochi, India (on line).
| | | Last update : 09/12/2020 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed April 01, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
