|
|
 |
| Speleophriidae Boxshall & Jaume, 2000 ( Misophrioida ) | | Ref.: | Boxshall & Halsey, 2004 (p.40; 221: Def.; p.222: Genera key); Vives & Shmeleva, 2010 (p.140, Rem., Genera key); Bradford-Grieve & al., 2014 (p.531: Rem.) | | Rem.: | Boxshall & Jaume (2000 a, p.1) créent une famille pour des genres initialement classés parmi les Misophriidae (see in Huys, 1988 a, p.150 & following). 7 G: Archimisophria, Boxshallia, Expansophria, Huysia, Protospeleophria, Speleophria, Speleophriopsis, (? plus 1 uncertain status: Dimisophria). |  issued from : D. Jaume, G.A. Boxshall & T.M. Iliffe in J. Nat. Hist., 1998, 32. [p.672, Table 1] Main distinguishing features between genera in the Spelepphria group. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity, Part II, The Ray Society, 2004, Part II, No 166. [p.223, Fig.58]. A, Expansophria galapagensis male; B, Speleophria bunderae female; C, Archimisophria discoveryi female urosome; D, Archimisophria squamosa P5 female; E, Boxshallia bulbantennulata A2; F, Mxp; G, female P5 and intercoxal sclerite.
From Boxshall & Iliffe, 1990: A; Jaume, Boxshall & Humphreys, 2001: B; Huys & Boxshall, 1991: C-G. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity, Part II, The Ray Society, 2004, Part II, No 166. [p.221]. Armature formula of swimming legs P1 to P4 and legs P5 male (m) and female (f). Nota: Swimming legs P1 to P4 biramous, typically with 3-segmented rami except endopod of P1 often 2-segmented due to failure of 2nd and 3rd segmets to separate; members of each leg pair joined by intercoxal sclerite; inner coxal seta and outer seta on basis present, inner seta on basis on P1 only. - Setation often reduced or modified by transformation of setae into spines. - P5 primitively biramous; members of leg pair joined by intercoxal sclerite; praecoxa usually absent; coxa and basis separate or fused; basis with outer seta; inner coxal seta absent; exopod typically 2-segmented and bearing up to 7 setal elements in female; 2 or 3-segmented and with up to 8 setae in male; endopod 1-segmented and bearing 1 seta in both sexes, or reduced to single seta as in Boxshallia. P5 usually uniramous due to loss of endopod; entire P5 absent in Expansophria apoda Boxshall & Iliffe, 1987. - P6 confluent in female, separate in male, represented by opercular plate closing off genital openings. | | | | | (1) Archimisophria Boxshall, 1983 | |
| | Ref.: | Boxshall, 1989 (p.522); Huys & Boxshall, 1991 (p.88, 407, 460); Boxshall & Halsey, 2004 (p.222) | | Rem.: | Total: 2 spp. | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 1.367 mm (n = 3; SD = 0.2517) and the mean male size is 1.20 mm (n = 2). The size ratio (male/female) is 0.94, or approximately 88-96 %. The sex ratio (female/male) is up to date 2/1 in 2014. | | | | (2) Boxshallia Huys, 1988 | |
| | Ref.: | Huys, 1988 a (p.139, Rem.); Boxshall, 1989 (p.522, 524); Huys & Boxshall, 1991 (p.88, 461); Jaume & al., 1998 (p.672); Boxshall & Halsey, 2004 (p.222); Vives & Shmeleva, 2010 (p.152, Rem.) | | Rem.: | Total: 1 sp. | | | | (0) Dimisophria Boxshall & Iliffe, 1987 | |
| | Ref.: | Boxshall & Iliffe, 1987 (p.245); Boxshall, 1989 (p.522); Huys & Boxshall, 1991 (p.88); Boxshall & Halsey, 2004 (p.222, Rem.); Vives & Shmeleva, 2010 (p.154, Rem.) | | Rem.: | type: Dimisophria cavernicola . Total: 1 sp. | | | | (3) Expansophria Boxshall & Iliffe, 1987 | |
| | Ref.: | Boxshall & Iliffe, 1987 (p.230); Boxshall, 1989 (p.522); Huys & Boxshall, 1991 (p.88); Jaume & Boxshall, 1996 a (p.1591, Rem: biogeography); Vives & Shmeleva, 2010 (p.141, Rem., spp. key) | | Rem.: | type: Expansophria dimorpha. Total 4 spp. | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 0.583 mm (n = 6; SD = 0.0698) and the mean male size is 0.48 mm (n = 2). The size ratio (male/female) is 0.84, or approximately 82-84 %. The sex ratio (female/male) is up to date 3/1 in 2014. | 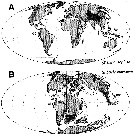 issued from : D. Jaume & G.A. Boxshall in J. Nat. Hist., 1996, 30. [p.1593, Fig.5]. A, The current known distribution of Expansophria and Speleophriopsis; B, same stations translated on the 120 Myr BP paleocoastline map (Maps adapted from A.G. Smith & al., 1994). According to the authors, the most parsimonious interpretation of the current known distribution of Expansophria (as Speleophriopsis) appears to be a relicts of an ancient (late Mesozoic), once widely distributed tropical/subtropical shallox-water fauna. Their known localities on a late Mesozoic paleocoastline map reveals a remarkable continuity between localities which are today separated by large distances and by continental land-masses. The coincidence with the borders of the former Tethys Sea is remarkable. The distribution of both taxa appears to be the result of a vicariant processes associated with tectonic plate motion, with the opening of the Atlantic (ca. 120Myr BP) and the closure of the Tethys Sea (ca. 20 Myr BP) as the major vicariant events.. This conforms exactly with the explanation currently offered for similarity disjunct modern distributions displayed by numerous stygobiont malacostracean genera with limited dispersal abilities (see in Stock, 1993). The presence of this genus (as Speleophriopsis in Mediterranean waters is especially notesworthy, as it apparently contradicts the widely held view that this sea does not harbour an ancient marine fauna. Such a fauna would have vanished during the Messinian Salinity Crisis, a putative complete desiccation event (approximately 5.5 Myr BP). Both taxa are tightly associated with coastal, hypogean waters and their potential for dispersal is limited. These characteristics indicate that it is unlikely that they could have invaded the Mediterranean Sea only after its reflooding during the Pliocene. These taxa inhabit waters with salinities only marginally reduced from the normal marine waters and never penetrate into brackish or fresh inland waters | | | | | (4) Huysia Jaume, Boxshall & Iliffe, 1998 | |
| | Ref.: | Jaume & al., 1998 (p.663); Boxshall & Halsey, 2004 (p.222) | | Rem.: | Type: Huysia bahamensis . Total: 1 sp. | | | | (5) Protospeleophria Jaume, Boxshall & Iliffe, 1998 | |
| | Ref.: | Jaume & al., 1998 (p.672); Boxshall & Halsey, 2004 (p.222) | | Rem.: | Total: 1 sp.: | | | | (6) Speleophria Boxshall & Iliffe,1986; emend. Jaume & Boxshall, 1996 | |
| | Ref.: | Boxshall & Iliffe, 1986 (p.55); Boxshall, 1989 (p.522); Huys & Boxshall, 1991 (p.88, 467); Jaume & Boxshall, 1996 (p.998, 1004, Rem.); Jaume & al., 2001 (p.20, Rem.); Boxshall & Halsey, 2004 (p.222); Karanovic & Eberharde, 2009 (p.61, spp. Key Females); Vives & Shmeleva, 2010 (p.144, Rem., spp. Key) | | Rem.: | Type: Speleohria bivexilla. Total: 5 spp. | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 0.556 mm (n = 9; SD = 0.1106) and the mean male size is 0.53 mm (n = 4, SD = 0.0658). The size ratio (male/female) is variable according to the species: 0.97- 0.74, or approximately 89 %. The sex ratio (female/male) is up to date 4/2 in 2014. | 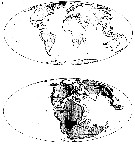 issued from : D. Jaume, G.A. Boxshall & W.F. Humphreys in Zool. J. Linnean Soc., 2001, 133. [p.21, Fig.14]. A: current known distribution of the calanoid genus Speleophria (black triangle) and the stygocyclopid Stygocyclopia 'black circle). B: same stations translated on a 155 Myr BP palaeocoastline map. (Maps adapted from A.G. Smith & al., 1994). | | | | | (7) Speleophriopsis Jaume & Boxshall, 1996 | |
| | Syn.: | Speleophria Boxshall & Iliffe, 1986 (part.) | | Ref.: | Jaume & Boxshall, 1996 (p.990, 1004, Rem.); 1996 a (p.1591, : biogeography); Boxshall & Halsey, 2004 (p.222); Vives & Shmeleva, 2010 (p.43, Rem., spp. Key) | | Rem.: | type: Speleophriopsis balearicus. Total: 4 spp. | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 0.720 mm (n = 4; SD = 0.2544) and the mean male size is 0.85 mm (n = 2). The size ratio (male/female) is variable according to the species 0.89-1.07, or approximately 98 %. The sex ratio (female/male) is up to date 4/2 in 2014. | issued from : D. Jaume & G.A. Boxshall in J. Nat. Hist., 1996, 30. [p.1593, Fig.5]. A, The current known distribution of Speleophriopsis and Expansophria; B, same stations translated on the 120 Myr BP paleocoastline map (Maps taken from Smith & al., 1994). According to the authors, the most parsimonious interpretation of the current known distribution of Speleophriopsis (as Expansophria) appears to be a relicts of an ancient (late Mesozoic), once widely distributed tropical/subtropical shallox-water fauna. Their known localities on a late Mesozoic paleocoastline map reveals a remarkable continuity between localities which are today separated by large distances and by continental land-masses. The coincidence with the borders of the former Tethys Sea is remarkable. The distribution of both taxa appears to be the result of a vicariant processes associated with tectonic plate motion, with the opening of the Atlantic (ca. 120 Myr BP) and the closure of the Tethys Sea (ca. 20 Myr BP) as the major vicariant events.. This conforms exactly with the explanation currently offered for similarity disjunct modern distributions displayed by numerous stygobiont malacostracean genera with limited dispersal abilities (see in Stock, 1993). The presence of this genus (as Expansophria in Mediterranean waters is especially notesworthy, as it apparently contradicts the widely held view that this sea does not harbour an ancient marine fauna. Such a fauna would have vanished during the Messinian Salinity Crisis, a putative complete desiccation event (approximately 5.5 Myr BP). Both taxa are tightly associated with coastal, hypogean waters and their potential for dispersal is limited. These characteristics indicate that it is unlikely that they could have invaded the Mediterranean Sea only after its reflooding during the Pliocene. These taxa inhabit waters with salinities only marginally reduced from the normal marine waters and never penetrate into brackish or fresh inland waters. See Discussion in Krsinic (2017, p.946) concerning the question of the Tethyan relicts. | | | | | | | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 25 mars 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
