|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Calanoidea ( Superfamille ) |
|
|
|
Paracalanidae ( Famille ) |
|
|
|
Paracalanus ( Genre ) |
|
|
| |
Paracalanus quasimodo Bowman, 1971 (F,M) | |
| | | | | | | Syn.: | ? Paracalanus parvus s.l. : Chihara & Murano, 1997 (p.845, Pl.135: F,M);
?Paracalanus sp. : Liang & Uye, 1996 (p.219, Rem.). | | | | Ref.: | | | Bowman, 1971 b (p.24, figs.F,M); Andronov, 1977 (p.155, fig.F); Bradford, 1978 (p.134: Rem.); Björnberg & al., 1981 (p.622, figs.F,M); Campaner, 1985 (p.10); Hiromi, 1987 (p.147, 153, 155, Rem.); Nishida, 1989 (p.173, table 1, 2, dorsal hump); Kang, 1996 (p.409, Rem.); Bradford-Grieve & al., 1999 (p.878, 910, figs.F,M); Kesarkar & Anil, 2010 (Rem.: p.406, fig.F) |  issued from : T.E. Bowman in Smithson. Contr. Zool., 1971, 96. [p.25, Fig.20]. Female (from 33°34'N, 78°24'W): a-b, habitus (lateral and dorsal, respectively); c, 5th pedigerous and genital segments (lateral); d-e-f, last thoracic segment, genital and 2nd urosomal segments (dorsal, right and left sides, respectively); g, anal segment and caudal rami (dorsal); h-k, P1 to P4, respectively; l, P5 Nota: Prosome about 3 times as long as urosome, with characteristic dorsal hump (similar to those in some species of Acrocalanus but less strongly developed). Head and 1st pedigerous somite fused, 4th and 5th separated by indistinct suture. Genital segment rather broad in relation to length; lateral part of posterior margin sometimes armed with row of minute spinules, lateral surface with cluster of spinules on either side, anterior and dorsal to spermatheca. Spermatheca obovate; remaining urosomites relatively broader and shorter than those of P. indicus. A1 25-segmented, reaching to about posterior margin of caudal ramus. Swimming legs pattern very similar to that of P. indicus , basipod conspicuously spinose. P5, terminal spine longer than 2nd segment.
|
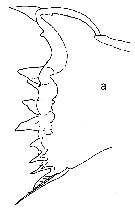 issued from : T.E. Bowman in Smithson. Contr. Zool., 1971, 96. [p.26, Fig.21, a]. Female: a, Md (cutting edge of mandibular blade).
|
 issued from : T.E. Bowman in Smithson. Contr. Zool., 1971, 96. [p.26, Fig.21, b-h]. Male: b, d, habitus (dorsal and lateral, respectively); c, forehead with base of A1 (lateral); e, anal segment and cuadal rami (dorsal); f, distal segments of A1; g, P5 (anterior); h, left P5 (lateral view).
|
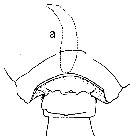 issued from : T.E. Bowman in Smithson. Contr. Zool., 1971, 96. [p.27, Fig.22, a]. Male: a, posterior prosome and genital segment (dorsal). Nota: Genital segment with cluster of spinules on either side, orherwise identical with P. indicus.
|
 issued from : K.S. Kesarkar & A.C. Anil in J. mar. Biol. Ass. UK, 2010, 90 (2); [p.406, Table 2]. Characteristics of females.
|
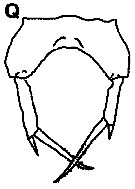 issued from : K.S. Kesarkar & A.C. Anil in J. mar. Biol. Ass. UK, 2010, 90 (2); [p.407, Fig.5, Q]. Female: Q, P5.
| | | | | Ref. compl.: | | | Björnberg, 1973 (p.304: Rem., 388); Kimmerer, 1993 (p.267, tab.2); Suarez Morales, 1994 b (tab.1); Eskinazi-Sant'Anna & Tundisi, 1996 (t.1,2); Suarez-Morales & Gasca, 1997 (p.1525); Bonecker & al., 1998 (p.31); Lopes & al., 1998 (p.195); Alvarez-Cadena & al., 1998 (tab.1,3); Mauchline, 1998 (tab. 62, 63); Suarez-Morales & Gasca, 1998 a (p.110); Lopes & al., 1999 (p.215, tab.1); Paffenhöfer & al., 1996 (p.1699, motion behavior); Neumann-Leitao & al., 1999 (p.153, tab.2); Ara, 2001 b (p.121); Dias & Bonecker, 2005 (p.100 + poster); Choi & al., 2005 (p.710: Tab.III) ; Araujo, 2006 (p.173, Tab.3): Sterza & Fernades, 2006 (p.95, Table 1, occurrence); Dias & Araujo, 2006 (p.64, Rem., chart); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Galbraith, 2009 (pers. comm.); Miyashita & al., 2009 (p.815, Tabl.II); Magalhaes & al., 2009 (p.187, Table 1, %); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Magris & al., 2011 (p.260, abundance, interannual variability); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Costa R.G. da & al., 2011 (p.364, Table 1, seasonal occurrence); Pinheiro & al., 2011 (p.425, abundance vs season); Almeida LR. & al., 2012 (p.13, Table 1, abundance); Miyashita & al., 2012 (p.1557, Table 2: occurrence); Gubanova & al., 2013 (in press, p.4, Table 2); Cornils & Blanco-Bercial, 2013 (p.861, Table 1, molecular analysis, figs.3, 4, 5); Garbosa da Costa & al., 2013 (p.756, Table 1, abundance vs tide); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production, Table 4: % vs. season, fig.8); Araujo & al., 2016 (p.1, Table 3, abundance, %) ; Rosa J.C.L. & al., 2016 (p.67, abundance, feeding, Rem.: p.71); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1, Rem.: p.502, abundance); Atique & al., 2017 (p.1, Table 1, fig.8, Rem.: p./11); Dias & al., 2018 (p.1, Tables 2, 5: vertical distribution, abundance vs. season); Dias & al., 2018 a (p.189, Rem.: p.196). | | | | NZ: | 5 + 1 douteuse | | |
|
Carte de distribution de Paracalanus quasimodo par zones géographiques
|
| | | | | |  issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.64]. issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.64].
Chart of occurrence in Brazilian waters (sampling between 22°-23° S).
Nota: 372 specimens. |
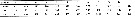 Issued from : J. da Cruz Lopes de Rosa, W.M. Monteiro-Ribas & L. Dias de AlmeidaFernandes in Br. J. Oceanogr., 2016, 64 (1). [p.70, Table 2]. Issued from : J. da Cruz Lopes de Rosa, W.M. Monteiro-Ribas & L. Dias de AlmeidaFernandes in Br. J. Oceanogr., 2016, 64 (1). [p.70, Table 2].
Paracalanus quasimodo density (inds. m3).
Samples collected monthly on days of full moon at Cabo Frio, Brazil (23.8° S, 42.018 °W) from January to December 2011, by horizontal hauls (1 m depth).
The dominance of P. quasimodo was due to its emphasis on herbivory (R2 = 0.83) and lead to a decrease in the planktonic diversity (R2 = 0.63).
The results revealed that predation by carnivorous copepods , like Corycaeus spp. and Oncaea spp, is the main opposing factor affecting P. quasimodo dominance (R2 = 0.70 |
| | | | Loc: | | | Brazil (Paranagua Bay, off Rio de Janeiro, Guanabara Bay, Campos Basin, Canaeia Lagoon, Mucuri estuary, Cabo Frio, Vitoria Bay, Vitoria - Cabo de Sao Tomé, Camamu, Guarairas Lagoon Recife, off Natal, Curuça estuary, Caeté Estuary, Amazon Estuary, Ajuruteua Beach), Caribbean Colombia, Bahia de Mochima (Venezuela), Caribbean Sea (Chetumal Bay), Yucatan (Ascension Bay), G. of Mexico, Florida, New York (in Judkins & al., 1980, p.674, as P. quasimodi), Chile (Björnberg, 1973; Rosales & Sepulveda, 1992: Valparaiso), ? Japan (cf. Uye, 1991, p.627; Uye & Shibuno, 1994; Liang & Uye, 1996), Oregon, California: Tomales Bay (in Kimmerer, 1993), Medit. (off W Jalitha Is., Black Sea)
Data from Cornils & Blanco-Bercial (2013): 37°30'N; 8°10'W | | | | N: | 41 | | | | Lg.: | | | (436) F: 1; M: ?; (202) F: 0,82-0,75; M: 0,82; {F: 0,75-1,00; M: 0,82}
The mean female size is 0.857 (n = 3; SD = 0.1290) and the mean male size near of 0.82. The size ratio (male : female) is more or less of 0.957. The sex ratio (female: male) is 1. | | | | Rem.: | Signalé par erreur en Méditerranée occidentale in Ounissi & Khelifi-Touhami (1999, p.10).
Certaines confusions sont possibles entre cette espèce et P. parvus ou P. indicus.
La présence de cette espèce à San Francisco pourrait avoir été introduite par les ballasts des navires, comme dans les eaux sino-japonaises.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 12/02/2021 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 27 mars 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)

;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
