|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Clausocalanoidea ( Superfamille ) |
|
|
|
Phaennidae ( Famille ) |
|
|
|
Phaenna ( Genre ) |
|
|
| |
Phaenna spinifera Claus, 1863 (F,M) | |
| | | | | | | Syn.: | ? Onchocalanus latus Esterly, 1911 (p.326, figs.F); 1912 (tab.10); 1913 (p.183, figs.F); Brodsky, 1950 (1967) (p.235, figs.F);
Onchocalanus nudipes C.B. Wilson, 1942 a (p.199, Descr.F, figs.F, [fig.83: juv.M); 1950 (p.273); Fagetti, 1962 (p.23) | | | | Ref.: | | | Claus, 1863 (p.189, figs.F,M); Giesbrecht, 1892 (p.293, 774, figs.F,M); Thompson, 1888 d (p.142); T. Scott, 1894 b (p.81, figs.M); Giesbrecht & Schmeil, 1898 (p.50, Rem. F,M); Thompson & Scott, 1903 (p.234, 246); Wolfenden, 1904 (p.111, 112); Farran, 1908 b (p.47) ; A. Scott, 1909 (p.80); Wolfenden, 1911 (p.285); With, 1915 (p.241, figs.F, Rem.F,M); Pesta, 1920 (p.514); Farran, 1926 (p.268); 1929 (p.209, 255); Sewell, 1929 (p.175); Rose, 1933 a (p.126, figs.F,M); Mori, 1937 (1964) (p.54, figs.F); Lysholm & al., 1945 (p.24); Vervoort, 1950 b (p.83, figs.F,M, juv.M, Rem.); Chiba, 1956 (p.32, figs.juv.F); Tanaka, 1960 a (p.85, figs. juv.F); Grice, 1962 (p.203, fig.F); Chen & Zhang, 1965 (p.57, figs.F,M); Vervoort, 1965 (p.25, Rem.); Owre & Foyo, 1967 (p.57, figs.F,M); Mazza, 1967 (p.174, figs.F,M, juv.); Park, 1968 (p.554); Vidal, 1968 (p.30, figs.F); Corral Estrada, 1970 (p.156; Bradford, 1970 a (p.355, figs.M); 1972 (p.40, fig.F); Razouls, 1972 (p.94, Annexe: p.52, figs.F,M); Bradford, 1973 (p.137, Rem.F,M, fig.2a); Marques, 1974 (p.15); 1976 (p.991); Björnberg & al., 1981 (p.635, figs.F,M); Zheng & al., 1982 (p.40, figs.F); Park, 1983 a (p.318, figs.F); Bradford & al., 1983 (p.70, figs.F,M); Roe, 1984 (p.357); Tanaka & Omori, 1992 (p.268); Mazzocchi & al., 1995 (p.188, figs.F, Rem.); Chihara & Murano, 1997 (p.854, Pl.137: F); Bradford-Grieve & al., 1999 (p.880, 928, 929, figs.F,M); Conway & al., 2003 (p.188, figs.F,M, Rem.); Boxshall & Halsey, 2004 (p.159, figs.F); Mulyadi, 2004 (p.187, figs.F, Rem.);Vives & Shmeleva, 2007 (p.687, figs.F,M, Rem.); Blanco-Bercial & al., 2011 (p.1, Table 103, molecular biology, phylogeny); Soh & al; (p.74, figs.F); Laakmann & al., 2019 (p.330, fig. 2, 3, phylogenetic relationships) | 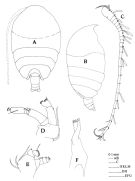 issued from : J.M. Bradford, L. Haakonssen & J.B. Jillett in Mem. N.Z. Oceanogr. Inst., 1983, 90. [p.62, Fig.34]. Female: A, habitus (dorsal); B, idem (lateral right side); C, A1; D, A2; E, Md (mandibular palp); F, Md (cutting blade).
|
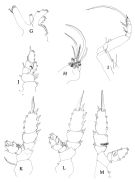 issued from : J.M. Bradford, L. Haakonssen & J.B. Jillett in Mem. N.Z. Oceanogr. Inst., 1983, 90. [p.63, Fig.34 (legend p.62)]. Female: G, Mx1; H, Mx2; I, Mxp; J, P1; K, P2; L, P3; M, P4.
|
 issued from : J.M. Bradford, L. Haakonssen & J.B. Jillett in Mem. N.Z. Oceanogr. Inst., 1983, 90. [p.64, Fig.35 (legend p.65)]. Male: A, habitus (dorsal); B, idem (lateral left side); C, A1; D, A2; E, Md (mandibular palp); F, Mx1; G, Mx2.
|
 issued from : J.M. Bradford, L. Haakonssen & J.B. Jillett in Mem. N.Z. Oceanogr. Inst., 1983, 90. [p.65, Fig.35]. Male: H, Mxp; I, P1; J, P2; K, P3; L, P4; M, P5.
|
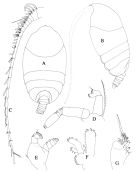
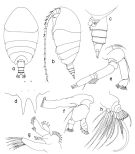 issued from : T. Park in Antarct. Res. Ser. Washington, 1983, 39 (5). [p.320, Fig.1]. Female (from southern coast of W Australia): a, habitus (dorsal); b, idem ( lateral left side); c, urosome (lateral left side); d, rostrum (anterior view); e, A2; f, Md; g, Mx1; h, Mx2. Nota: 1st matasomal segment fully separate from head, 4th and 5th only partially fused close to ventral margin.
|
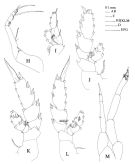
 issued from : T. Park in Antarct. Res. Ser. Washington, 1983, 39 (5). [p321, Fig.2]. Female: a, Mxp; b, P1 (anterior); c, P2 (posterior); d, P3 (posterior); e, P4 (posterior).
|
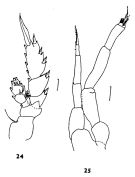 issued from : J.M. Bradford in N.Z. Jl Mar. Freshw. Res., 1970, 4 (4). [p.355, Figs 24-25]. Male (off Kaikoura, New Zealand): 24, P3; 25, P5. Scale bars represent 0.1 mm.
|
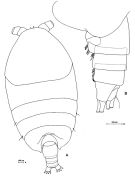 Issued from: M.G. Mazzocchi, G. Zagami, A. Ianora, L. Guglielmo & J. Hure in Atlas of Marine Zooplankton Straits of Magellan. Copepods. L. Guglielmo & A. Ianora (Eds.), 1995. [p.189, Fig.3.33.1]. Female: A, habitus (dorsal); B, urosome (lateral left side). Nota: head and 1st thoracic segment separated, 4th and 5th incompletely separated. Lateral thoracic somites 3 and 4 with a rather long sensillum on either side. Ventral surface of urosomal somites ornated with stout setae (this ornementation is fragile and can easily be lost in fixed specimens).
|
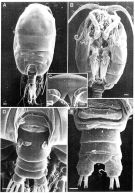 Issued from: M.G. Mazzocchi, G. Zagami, A. Ianora, L. Guglielmo & J. Hure in Atlas of Marine Zooplankton Straits of Magellan. Copepods. L. Guglielmo & A. Ianora (Eds.), 1995. [p.190, Fig.3.33.2]. Female (SEM preparation): A, habitus (dorsal); B, cephalothorax (ventral ); C, forehead (frontal view); D, urosome (dorsal); E, idem (ventral). Bars: A, B, D 0.100 mm; C, E 0.050 mm.
|
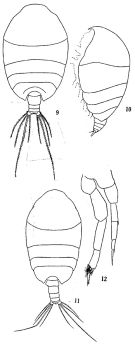 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.17, 9-12]. Female (from E China Sea): 9, habitus (dorsal); 10, idem (lateral left side). Male: 11, habitus (dorsal); 12, P5 (posterior).
|
 issued from : C.B. Wilson in Scientific Results of Cruise VII of the Carnegie during 1928-1929. Biology-I. Carnegie Inst. Wash. Publ. 536, 1942. [p.227, Fig.74]. As Onchocalanus nudipes. Female: 74, Mx2.
|
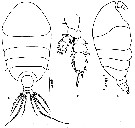 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.40, Fig.23]. Female: a-b, habitus (dorsal and lateral, respectively); c, P3. Scale bars in mm.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.27, Figs.9-15]. Female: 9, P2 (posterior); 10, P3 (posterior); 11, P4 (posterior); 12, Mx1; 13, P1; 14, habitus (dorsal); 15, A2.
|
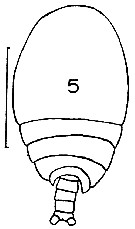 issued from : J.M. Bradford in Mem. N. Z. Oceonogr. Inst., 1972, 54. [p.42, Fig.9, (5)]. Female (from Kaikoura, New Zealand): 5, habitus (dorsal). Scale bar: 1mm.
|
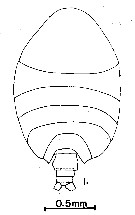 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.204, Pl.15, Fig.1]. Female (from equatorial Pacific): 1, habitus (doprsal). Nota: Genital segment with a large protrusion on the right side from which arises a smaller spine-like process.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.12, Figs.2, 3, 8, 36]. Female: 2, A2 (anterior surface); 3, Palp of Md (anterior surface); 8, Mxp (anterior surface); 36, Mx1 (posterior view).
|
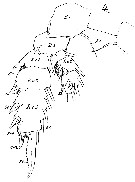 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.12, Fig.4]. Female: 4, P3 (posterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.12, Figs.5, 35]. Male: 5, P5 (posterior view); 35, Mx1 ( (anterior view). Ps = left leg; Pd = right leg.
|
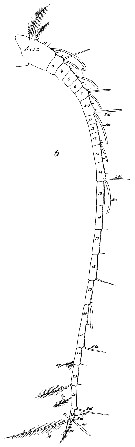
 issued from : C. With in The Danish Ingolf-Expedition, Copepoda I, 1915, III, 4. [p.242, Text-fig. 79]. Female (from 51°00N, 11°43'W): Lateral outline of body). Nota: In some of spexcimens the head possesses, dorsally, a conical protuberance. Length of prosome: 2.42 mm, urosome: 0.48 mm. The legs are scarcely different from Giesbrechts description. The outer margin of the 3rd exopodal segment of P1 has proximally to the middle a small incision and a glandular pore. On the anterior surface of the 3rd endoppodal segment of P2 and P3 a minute glandular pore, but in P4 none was seen. At the base of the outer spine in the exopodal segment 2, and at the base of the outer seta 1 and 3 of exopodal segment 3, glandular pores were found in P2, and in P3 and P4 at the base of the outer spine of exopodal 1. The endopodal segments 2-3, which are fused in P2, possess on the posterior surface two coronas of 8 and 6 long ; spines respectively; the endopodal segments 2 and 3 of P3 have 8 and 6 spines respectively ; the endopodal segment 2 of P4 has 6 long rather weak spines. Male: Length of prosome: 1.89 mm, urosome: 0.37 mm. The prosome did not possess a lamelliform projection, as figured by Giesbrecht; no dorsal protuberance.
|
 issued from : C. With in The Danish Ingolf-Expedition, Copepoda I, 1915, III, 4. [Pl. VII, Fig.1, a-c]. Female: a, rostrum (anterior view); b, labrum (oral view); c, lamina labialis and serrula 6-dentata. Nota: The very short labrum and the elevation in front of it are only indistinctly separated, and the latter is placed behind the insertion of the A1; the epistoma is covered by a number of longer and shorter setae; on the labrum proper a transverse group of fairly short setae; the marginal setae are only poorly developed. The oral surface (fig.b) (fig.b) has in front 4 longer and shorter oblique groups of shorter hairs converging towards the middle, and more posteriorly, probably composed of the 3 hinder, a longitudinally placed elongated group of short hairs; in the middle at least 3 groups of very short hairs or granules. The lamina labialis is only poorly developed (fig.c); in front of it a regular granular area; the area behind the lamina and on the labial lobes not studied in details.
|
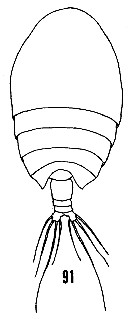 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1967, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. [p.23, Fig.91]. Female: 91, habitus (dorsal).
|
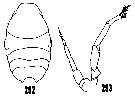 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1967, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. [p.50, Figs.292, 293]. After Sars, 1925 and Giesbrecht, 1892, respectively. Male: 292, prosome (dorsal); 293, P5.
|
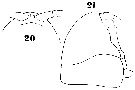 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.37, Figs.20, 21]. Female: 20, forehead (ventral); 21, same (lateral).
|
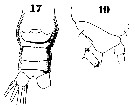 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.37, Figs.17, 19]. Female: 17, urosome (ventral); 19, thoracic segments 4-5 and urosomal segment 1-2 (lateral).
|
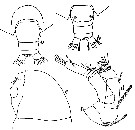
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.12, Fig.37]. Female: 37, Md (cutting edge of gnathobase).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.12, Fig.6]. Male: 6, A1 (ventral view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.12, Fig.1]. Male: 1, Mxp (anterior view).
|
 issued from : C.B. Wilson in Scientific Results of Cruise VII of the Carnegie during 1928-1929. Biology-I. Carnegie Inst. Wash. Publ., 1942, n° 536. [p.227, Figs.71-83]. As Onchocalanus nudipes. Female (from Pacific): 71, habitus (dorsal); 72, A2; 73, Mx1; 74, Mx2; 75, Mxp; 76, P1; 77, endopod of P1 (enlarged); 78, P1 from another specimen; 79, P2; 80, P2 from another specimen); 81, P3; 82, P4; 83, P5. Nota: Probably fig.83 is the P5 from juvenile male and an error in the legend (p.226), or an intersex form. After the author numerpous stages of development were taken in the eastern, central, and western parts of the tropical Pacific. No adult males were obtained, althrough some of the younger stages were probably undeveloped males. See in Vervoort's (1950 b, p.83) commentaries concerning the synonymy and the redescription of this species.
|
 issued from : W. Vervoort in Proc. K. ned. Akad. Wet., 1950, LIII, 1. [p.84, Fig.1]. Female (from ''Carnegie'' collection, 1°32'S, 82°16'W): a, habitus (dorsal view); b, urosome (lateral view from left side; c, left Md (mandibular palpus). Nota: Urosome very short, 4-segmented, the 4th (anal) segment completely telescoped into the preceding. Proportional length of segments and caudal rami 44 : 19 : 15 : 4 : 18 = 100. Urosomal segments with some scattered hair-like scales of characteristic appearance.
|
 issued from : W. Vervoort in Proc. K. ned. Akad. Wet., 1950, LIII, 1. [p.85, Fig.2]. Female: a, urosome (dorsal); b, same (ventral); c, head (lateral); d, A2 (posterior surface). Nota: Genital segment ventrally with a broadly rounded genital flap, covering the orifices of 2 small, scarcely visible receptacles. Rem.: After Vervvort (1950, p.86) Wilson's drawings of the A2, Mx1, Mx2 and Mxp all are very inaccurate with regards to the number of setae on the various lobes and segments.
|
 issued from : W. Vervoort in Proc. K. ned. Akad. Wet., 1950, LIII, 1. [p.85, Fig.3]. Female: a, left Md (manducatory plate, interior surface); b, left Mx1 (posterior surface); c, left Mxp (anterior surface). Nota: Mx1 small; 1st inner lobe elongated, carrying 6 strong, more or less plumose setiform spines and 4 smaller setae, one of which is very small and delicate; 2nd inner lobe with 1 single seta; 3rd inner lobe with 3 setae; 1st outer lobe with 9 strong, plumose setae; exopod with 5 setae; 2nd basal segment with 3 setae; endopod with 6 setae, the jointing between the endopodal joints is indistinct, but apparently there is 1 seta on the 1st, 1 on the 2nd and 4 on the 3rd (terminal ) joint.
|
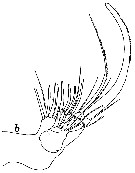 issued from : W. Vervoort in Proc. K. ned. Akad. Wet., 1950, LIII, 1. [p.87, Fig.4, b]. Female: b, right Mx2 (interior surface). Nota: Mx2 strongly developed. 1st basal segment carries 4 lobes, on the 1st lobe there are 4 setae of about equal length and 1 smaller seta, on the 2nd lobe there are 3 equal setae, on the 3rd 2 subequal setae, and on the 4th 1 strong, spinulose seta and a much smaller and shorter seta. The denticles on the last mentioned strong seta on the 4th lobe are minute. 2nd basal segment terminating into a single (5th) lobe, which carries a strong, curved, spiniform seta (this seta is much longer than the other setae, there are rather coarse denticles along the convex border, from the apex down to about half its length), in addition there are 2 muvch smaller setae on the 5th lobe. The segmentation of the endopod is very indistinct; there are 6 brush-shaped sensory appendages and 2 much longer, filiform appendages (the extremities of these sensory filaments are coverd by a mucous layer. There is no seta along the external margin of Mx2.
|
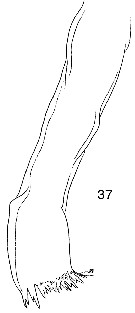
 issued from : W. Vervoort in Proc. K. ned. Akad. Wet., 1950, LIII, 1. [p.90, Fig.5]. Female: a, left P1; b, left P2; c, left P3; d, left P4 Of all legs the posterior surface has been figured. b, c, d at the same scale; a: magnified twofold from the others. After vervoort (1950, p.86) the figures of P3 and P4 apparently have been changed in Wilson's paper (fig.81 represents P4, fig.82 the P3.
|
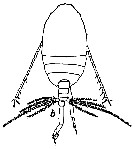 issued from : W. Vervoort in Proc. K. ned. Akad. Wet., 1950, LIII, 1. [p.90, Fig.6, b]. Male: b, adult habitus (dorsal aspect).
|
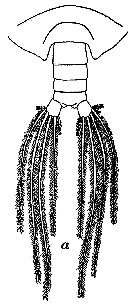 issued from : W. Vervoort in Proc. K. ned. Akad. Wet., 1950, LIII, 1. [p.90, Fig.6, a]. Male 5th copepodid stage: a, urosome (dorsal view).
|
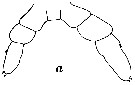 issued from : W. Vervoort in Proc. K. ned. Akad. Wet., 1950, LIII, 1. [p.87, Fig.4, a]. Male 5th copepodid stage: a, P5 (anterior surface).
|
 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.188, Fig.107]. Female (from East Indonesian Seas): a, habitus (dorsal); b, urosome (dorsal); c, P1; d, P2. Male: e, habitus (dorsal); f, P5. Nota Female: Exopod segment 2 armed with transverse rows of acicular spines on posterior surface. P5 absent.
|
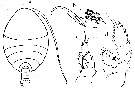 Issued from : O. Tanaka in Publ. Seto Mar. Biol. Lab., 1960, VIII (1). [p.85, Fig.1]. Immature female (from Sagami, middle Japan): a, habitus (dorsal); b, distal segments of Mx2; c, P1; d, P2. Nota: Body size = 1.81 mm.
|
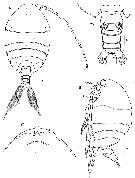 Issued from : H.Y. Soh, S.Y. Moon & J.H. Wi in Invertebrate Fauna of Korea (eds) Incheon: NIBR, 2013, 21 (28). [p.76, Fig.43]. Female (from Korean waters): A-B, habitus (dorsal and lateral, respectively); C, forehead (ventral); D, urosome (ventral). Scale bars: A, B = 400 µm; C = 200 µm; D = 100 µm.
|
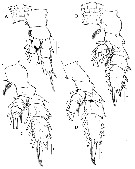 Issued from : H.Y. Soh, S.Y. Moon & J.H. Wi in Invertebrate Fauna of Korea (eds) Incheon: NIBR, 2013, 21 (28). [p.77, Fig.44]. Female: A, P1; B, P2; C, P3; D, P4. Scale bars: 100 µm.
| | | | | Ref. compl.: | | | Cleve, 1904 a (p.194); Pearson, 1906 (p.21, Rem.); Wilson, 1942 a (p.202); 1950 (p.285); Yamazi, 1958 (p.149, Rem.); Fagetti, 1962 (p.22); Gaudy, 1962 (p.93, 99, Rem.: p.107); Duran, 1963 (p.16); V.N. Greze, 1963 a (tabl.2); Gaudy, 1963 (p.23, Rem.); De Decker & Mombeck, 1964 (p.13); Shmeleva, 1965 b (p.1350, lengths-volume -weight relation); Mazza, 1966 (p.70); Pavlova, 1966 (p.43); Ehrhardt, 1967 (p.738, geographic distribution, Rem.); Fleminger, 1967 a (tabl.1); Delalo, 1968 (p.138); Evans, 1968 (p.14); Berdugo & Kimor, 1968 (p.448); Dowidar & El-Maghraby, 1970 (p.269); Morris, 1970 (p.2300); Deevey, 1971 (p.224); Bainbridge, 1972 (p.61, Appendix Table III: occurrence); Apostolopoulou, 1972 (p.327, 349); Binet & al., 1972 (p.55); Roe, 1972 (p.277, tabl.1, tabl.2); Björnberg, 1973 (p.330, 388); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Vives & al., 1975 (p.42, tab.II); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Carter, 1977 (1978) (p.35); Dessier, 1979 (p.205); Vaissière & Séguin, 1980 (p.23, tab.3); Porumb, 1980 (p.168); Kovalev & Shmeleva, 1982 (p.83); Vives, 1982 (p.291); Dessier, 1983 (p.89, Tableau 1, Rem., %); Turner & Dagg, 1983 (p.16, 22); De Decker, 1984 (p.316); Brenning, 1984 (p.3, Rem.); Scotto di Carlo & al., 1984 (p.1044); Cummings, 1984 (p.163, Table 2); Guangshan & Honglin, 1984 (p.118, tab.); Almeida Prado Por, 1985 (p.250); Brenning, 1985 a (p.28, Table 2); Brinton & al., 1986 (p.228, Table 1); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Madhupratap & Haridas, 1986 (p.105, tab.1); Jimenez-Perez & Lara-Lara, 1988; Lozano Soldevilla & al., 1988 (p.58); Hernandez-Trujillo, 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Heinrich, 1990 (p.17); Madhupratap & Haridas, 1990 (p.305, fig.5: vertical distribution night/day; fig.7: cluster); Suarez & al., 1990 (tab.2); Suarez & Gasca, 1991 (tab.2); Hernandez-Trujillo, 1991 (1993) (tab.I); Suarez, 1992 (App.1); Seguin & al., 1993 (p.23); Shih & Young, 1995 (p.71); Suarez-Morales & Gasca, 1997 (p.1525); Hure & Krsinic, 1998 (p.101); Gilabert & Moreno, 1998 (tab.1, 2); Suarez-Morales & Gasca, 1998 a (p.110); Smith S. & al., 1998 (p.2369, Table 6, moonsoon effects); Neumann-Leitao & al., 1999 (p.153, tab.2); Siokou-Frangou, 1999 (p.476); Lapernat, 2000 (tabl.3, 4); Razouls & al., 2000 (p.343, Appendix); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); Moraitou-Apostolopoulou & al., 2000 (tab.I); Lopez-Salgado & al., 2000 (tab.1); Holmes, 2001 (p.56, Rem.); Hidalgo & Escribano, 2001 (p.159, tab.2); Sameoto & al., 2002 (p.13); Beaugrand & al., 2002 (p.179, figs.5, 6); Vukanic, 2003 (139, tab.1); Hwang & al., 2003 (p.193, tab.2); Shimode & Shirayama, 2004 (tab.2); Hsiao & al., 2004 (p.326, tab.1); Gallienne & al., 2004 (p.5, tab.3); Lo & al., 2004 (p.89, tab.1); Zuo & al., 2006 (p.162: tab.1); Lavaniegos & Jiménez-Pérez, 2006 (p.151, tab.2, 4, Rem.); Dur & al., 2007 (p.197, Table IV); Khelifi-Touhami & al., 2007 (p.327, Table 1); Jitlang & al., 2008 (p.65, Table 1); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Cabal & al., 2008 (289, Table 1); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez Suarez-Morales, 2008 (p.521); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Galbraith, 2009 (pers. comm.); Williamson & McGowan, 2010 (p.273, Table III, Pacific central gyres: N & S); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Hidalgo & al., 2010 (p.2089, Table 2); Mazzocchi & Di Capua, 2010 (p.426); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Hsiao S.H. & al., 2011 (p.475, Appendix I); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Tutasi & al., 2011 (p.791, Table 2, abundance distribution vs La Niña event); Shiganova & al., 2012 (p.61, Table 4); Uysal & Shmeleva, 2012 (p.909, Table I); Salah S. & al., 2012 (p.155, Tableau 1); Dorgham & al., 2012 (p.473, Table 4: abundance vs season); Palomares-Garcia & al., 2013 (p.1009, Table I, V, abundance vs environmental factors); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Hirai & al., 2013 (p.1, Table I, molecular marker); Lidvanov & al., 2013 (p.290, Table 2, % composition); Zakaria & al., 2016 (p.1, Table 1); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence) ; El Arraj & al., 2017 (p.272, table 2, spatial distribution); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Belmonte, 2018 (p.273, Table I: Italian zones); Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence). | | | | NZ: | 21 | | |
|
Carte de distribution de Phaenna spinifera par zones géographiques
|
| | | | | | | | | | | | | | |  issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 22 ]. Phaenna spinifera (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 22 ]. Phaenna spinifera (from South Adriatic).
Dimensions, volume and Weight wet. Means for 50-60 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
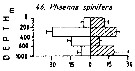 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.312, Fig.5]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.312, Fig.5].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 46- Phaenna spinifera.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
| | | | Loc: | | | Cosmopolite (tropical et temperate): South Africa (S & E), Atlant., G. of Guinea, Brazil (off Natal), Caribbean Sea, Caribbean Colombia, Yucatan, G. of Mexico, Florida, off Bermuda: Station "S" (32°10'N, 64°30'W), Long Island, off E Nova Scotia, G. of Guinea, off NW Cape Verde Is., off Morocco-Mauritania, Cap Ghir (Morocco), Canary Is., off Madeira, Azores, Bay of Biscay, off W Cabo Finisterre, off W Ireland, Ibero-moroccan Bay, Medit. (Alboran Sea, Sidi Fredj coast, Gulf of Annaba, Banyuls, G. of Lion, Marseille, Ligurian Sea, Tyrrhenian Sea, Napoli, Messina, Malta, Adriatic Sea, Ionian Sea, Aegean Sea, Lebanon Basin, W Egyptian coast, Alexandria, Black Sea), Sharm El-Sheikh, Safaga, Red Sea, Arabian Sea, Natal, Mascarene Basin, Rodrigues Is. - Seychelles, off SW Sri Lanka, Bay of Bengal, Flores Sea, Celebes Sea, Banda Sea, Ambon Bay (SW Ceram Is.), China Seas (East China Sea, South China Sea), Korea, Taiwan (SW, E, N: Mienhua Canyon), Japan (Tanabe Bay), Vancouver Is., Pacif. (W equatorial), Pacific (central gyres: N & S), BaJa California (Bahia Magdalena, W), Gulf of California, W Costa Rica, Bahia Cupica (Colombia), Galapagos-Ecuador, Peru, Chile (N-S, Mejillones Peninsula); Also sub-Antarct. (Pacif. SE), Straits of Magellan (Pacific area) | | | | N: | 190 | | | | Lg.: | | | (7) F: 2,9; M: 2,26; (9) F: 2,3; 2,05; M: 2,5; (15) F: 2,22-2,2; (16) F: 2,3-2,13; (35) F: 2,23; M: 2,1; (36) F: 2,88-3,02; (38) F: 2,34; M: 2,22; (46) F: 2,1-1,8; M: 1,8; (57) F: 2,25; (72) F: 2,4-1,84; M: 1,94; (101) F: 1,9-1,6; (116) F: 2,2; (180) F: 1,82; (187) F: 2,12; (199) F: 2,2-1,98; M: 1,98; (207) F: 1,94-1,89; (231) M: 2,5; (290) F: 1,9-2; M: 1,9-1,95; (313) M: 2,5; (333) F: 1,5; (335) F: 1,71; (447) F: 2,07; M: 1,89; (449) F: 2,1-1,8; M: 1,8; (867) F: 1,6-2,73; (991) F: 1,5-2,9; M: 1,8-2,5; (1023) F: 1,89; (1111) F: 2,16; (1122) F: 2,1; (1174) F: 2,45; {F: 1,50-3,02; M: 1,80-2,50} | | | | Rem.: | épi-mésopélagique. After Pearson (1906, p.21): vertical range down to 2900 m.
Sampling depth (sub-Antarct.) : 250 m.
With (1915) signale la présence dune petite protubérance dorsale sur le cephalosome de certains individus. Caractéristique observée également au large de la côte algérienne (Kerzabi, 1999, comm. pers.).
Voir aussi les remarques en anglais | | | Dernière mise à jour : 28/10/2022 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 14 juin 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
