|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Diaptomoidea ( Superfamille ) |
|
|
|
Pontellidae ( Famille ) |
|
|
|
Pontellina ( Genre ) |
|
|
| |
Pontellina plumata (Dana, 1849) (F,M) | |
| | | | | | | Syn.: | Pontella plumata Dana,1849; Brady, 1883 (part., p.92, figs.1-10); Kim & al., 1993 (p.270);
Calanops messinensis Claus, 1863 (p.212);
no Pontella plumata : Brady, 1883 (p.93, Pl.37, figs.11);
? Pontellopsis speciosus Brady, 1915;
? Pontellopsis aequalis Mori,1932 a (p.172, figs.M);
no Pontellina navalium Oliveira, 1946 (p.472); Vervoort, 1965 (p.191);
Pontella plumata : Webber & Roff, 1995 (tab.1);
Pontellina plumata s.l. : Brinton & al., 1986 (p.228, Table 1) | | | | Ref.: | | | Giesbrecht, 1892 (p.497, 774, figs.F,M); Thompson, 1888 d (p.143); Giesbrecht & Schmeil, 1898 (p.149, Rem.); Thompson & Scott, 1903 (p.236, 253); Wolfenden, 1905 (1906) (p.1022); A. Scott, 1909 (p.175, Rem.); Wolfenden, 1911 (p.362); Sewell, 1912 (p.354); 1914 a (p.240); Sars, 1925 (p.355); Farran, 1929 (p.210, 280); Rose, 1929 (p.46); Sewell, 1932 (p.391); Wilson, 1932 a (p.156, figs.F,M); Rose, 1933 a (p.265, figs.F,M); Farran, 1936 a (p.118); Mori, 1937 (1964) (p.100, figs.F,M); Dakin & Colefax, 1940 (p.99, figs.F); Wilson, 1942 a (p.204, fig.M); Chiba, 1956 (p.61, figs.F); Lysholm & al., 1945 (p.42); Sewell, 1947 (p.251); Chiba & al., 1957 a (p.12); Fish, 1962 (p.19); Brodsky, 1962 c (p.147, figs.M); Voronina, 1962 a (p.68); Grice, 1962 (p.240, figs.F,M); Gaudy, 1963 (p.29); Paiva, 1963 (p.74); Kasturirangan, 1963 (p.59, figs.F,M); Tanaka, 1964 c (p.270); Chen & Zhang, 1965 (p.109, figs.F,M); Vervoort, 1965 (p.190, Rem.); Saraswathy, 1966 (1967) (p.84); Owre & Foyo, 1967 (p.99, figs.F,M); Vidal, 1968 (p.45, figs.F); Park, 1968 (p.569, figs.F, Rem.); Corral Estrada, 1970 (p.204, Rem., figs.F,M); Ramirez, 1971 (p.88); Shih & al., 1971 (p.44); Silas & Pillai, 1973 (1974) (p.847, figs.F,M, Rem.); Marques, 1973 (p.245, figs.F); 1974 (p.18); Fleminger & Hulsemann, 1974 (p.71, figs.F,M); Fleminger, 1975 (p.395); Hulsemann & Fleminger, 1975 (p.174, figs.Juv. V & F,M), Rem.: development); Silas & Pillai, 1976 (p.847, Rem., figs.F,M); Marques, 1976 (p.996); Björnberg & al., 1981 (p.659, figs.F,M); Marques, 1982 (p.765); Chahsavar-Archad & Razouls, 1982 (p.32, figs.F); Lakkis, 1984 (p.297); Zheng Zhong & al., 1984 (1989) (p.257, figs.F,M); Kimmerer & al., 1985 (p.428); Hulsemann & Fleminger, 1990 (p.99, figs.F); Ohtsuka & Onbé, 1991 (p.214); Ianora & al., 1992 (p.401, fig.); Chihara & Murano, 1997 (p.873, Pl.161: F,M); Bradford-Grieve & al., 1999 (p.885, 960, figs.F,M); Bradford-Grieve, 1999 b (p.205, figs.F,M, Rem., figs.186, 194); Mulyadi, 2002 (p.157, figs.F,M, Rem.); Conway & al., 2003 (p.135, figs.F,M, Rem.).; G. Harding, 2004 (p.16, figs.F,M); Vives & Shmeleva, 2007 (p.516, figs.F,M, Rem.); Phukham, 2008 (p.100, figs.F,M); Blanco-Bercial & al., 2011 (p.1, Table 103, Biol. mol, phylogeny) | 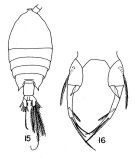 Issued from : T.S. Park in Fishery Bull. Fish Wild. Serv. U.S., 1968, 66 (3). [p.568, Pl.13, Figs.15-16]. Female: 15, habitus (dorsal); 16, P5. Nota: The right and left endopods are slightly unequal, and each is single-pointed, not forked.
|
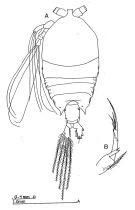 issued from : Bradford-Grieve J.M. in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). NIWA Biodiversity Memoir, 111, 1999. [p.206, Fig.153]. Female: A, habitus (dorsal); B, P5. Nota: The endopod shows the distal part forked contrary to Park (1968).
|
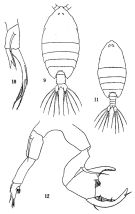 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.48, 9-12]. Female (from E China Sea): 9, habitus (dorsal); 10, right P5 (posterior). Male: 11, habitus (dorsal); 12, P5 (posterior).
|
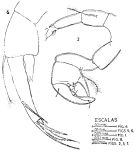
 issued from : F.C. Ramirez in Revta Mus. La Plata, Seccion Zool., 1971, XI. [Lam.II, figs.2, 8]. Female (from off Mar del Plata): 2, P5; 8, habitus (dorsal). Scale bars in mm: 0.05 (2); 0.5 (8).
|
 issued from : J. Corral Estrada in Tesis Doct., Univ. Madrid, A-129, Sec. Biologicas, 1970. [Lam.54, figs.1-2]. Female (from Canarias Is.): 1, P5. Male: 2, P5.
|
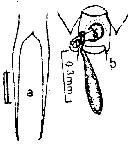 issued from : E.G. Silas & P.P. Pillai in J. mar. biol. Ass. India, 1973 (1976), 15 (2). [p.849, Fig.35, a-b]. Female (from Indian Ocean): a, rostrum (anteriorl view); b, genital segment with spermatophore (ventral). Scale as in Calanopia minor.
|
 issued from : E.G. Silas & P.P. Pillai in J. mar. biol. Ass. India, 1973 (1976), 15 (2). [p.848, Fig.34]. Female: a, urosome (dorsal); b, P5. Male: c, A1; d, P5. Scale as in Calanopia minor.
|
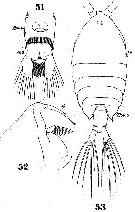 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.40, Figs.51, 52, 53]. Female: 51, urosome (ventral); 52, forehead (lateral); 53, habitus (dorsal).
|
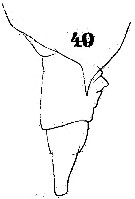 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.40, Fig.49]. Female: 49, Th5 and urosome (lateral).
|
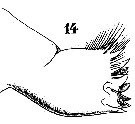 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Fig.14]. Female: 14, Md (mandibular blade, postrior view).
|
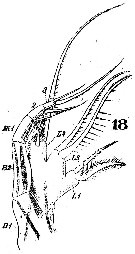 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Fig.18]. Female: 18, Mxp (anterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Fig.36]. Female: 36, P5 (anterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Fig.4]; Male: 4, right A1 (ventral view).
|
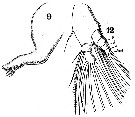 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Figs.9, 12]. Male: 9, Md (mandibular blade); 12, Md (mandibular palp, anterior view).
|
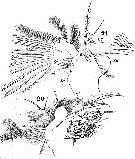 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Figs.20, 21]. Male: 20, Mx1 (posterior view); 21, idem (anterior view).
|
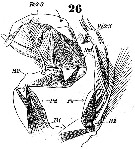 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.25, Fig.26]. Male: 26, P5 (posterior view).
|
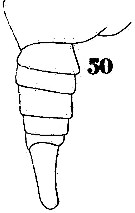 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.40, Fig.50]. Male: 50, Th5 and urosome (lateral).
|
 issued from : A. Fleminger & K. Hulsemann in Fishery bulletin, 1974, 72 (1). [p.72, Fig.3]. Female (from different seas): a, Th 4-5 and genital segment; b, habitus (lateral right side; other specimen); c, rostral filaments; d, range of variation observed in Th 4-5 (lateral view); e, Th 4-5 and urosome (dorsal; same specimen as a); f, range of variation observed in Th 4-5 (dorsal); g, habitus (dorsal; same specimen as b); h, lateral margin of right caudal ramus of various specimens (dorsal). Nota: Right caudal ramus fused to anal segment, lateral margin with small pointed projection somewhat variable in shape and size just anterior to base of outermost seta, ratio of length to width: median 1.53, range = 1.28-1.74:1, showing apparent overlap only with the equatorial Atlantic congeners; left caudal ramus not fused and longer than right one. P5 with inner margin of exopod lacking hair, endopod polymorphic with 1 or 2 apical spines fused to segment, spines on left and right endopod may differ in number in same specimen; exopod 2 to 3 times longer than endopod (median: 2.37:1, range = 1.97-3.08:1), differing strongly from Indian and Pacific congeners.
|
 issued from : A. Fleminger & K. Hulsemann in Fishery bulletin, 1974, 72 (1). [p.74, Fig.4]. Female: a, P5 (anterior view); b, endopod of P5 of other specimens; c, enlargement of P5 apex; d, enlargement of distal process endopod P5. Male: e, habitus (lateral right side); f, Th 4-5 and part of urosome and P5 (lateral right side); g, variation observed in Th 4-5 (lateral view); h, habitus (dorsal; same as e); i, P5 (posterior view; same as f); j, P5 chela (lateral view; same as f); k, Md (masticatory edge of the gnathobase; lateral view). Nota: Right caudal ramus not fused to anal segment, about equal in length to left ramus; ratio of length to width: median = 2.30:1, range = 2.0-2.53:1, but showing more overlap with congeners than found among females. Left P5 with proximal segment of exopod short relative to other congeners.
|
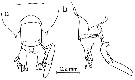 issued from : A. Fleminger & K. Hulsemann in Fishery bulletin, 1974, 72 (1). [p.101, Fig.33, a-b]. Female: Th 4-5 and urosome with attached spermatophore (dorsal and lateral respectively).
|
 issued from : A. Fleminger & K. Hulsemann in Fishery bulletin, 1974, 72 (1). [p.111, Table 19]. List of identified particles from microscopic analysis of stomach contents in adulte female.
|
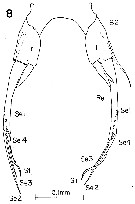 issued from : K. Hulsemann & A. Fleminger in Bull. Mar. Sci., 1975, 25 (2). [p.178, Fig.8]; Female: P5. l = left leg; r = right leg; B2 = 2nd basal segment; Re = exopod, Se 1-4 = 1st to 4th seta, Si = medial seta, St = terminal seta.
|
 issued from : K. Hulsemann & A. Fleminger in Bull. Mar. Sci., 1975, 25 (2). [p.181, Figs.16-17]. Male: 16, P5 (posterior view); 17, chela of chela in lateral view).
|
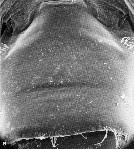 issued from : K. Hulsemann & A. Fleminger in Mar. Biol., 1990, 105. [p.101, Fig.1, a]. Female: a, genital segment (dorsal view). Nota: Scanning electron microscope examination revealed on the genital segment many patches of small spinules ornamenting the integument dorsally and laterally. The spinules are flattened, triangular, non-articulated, rigid structures rising from the integument, usually pointing obliquely posteriad. Nota: The adaptative value of spinules in Pontellina spp. remains obscure. Position and mutually arrangment of the prominent lateral clusters on the genital segment may be construed as functioning during courtship, by providing a specialized site for the male to grasp or stroke. Blades & Youngbluth (1979) report such activity by male Labidocera aestiva prior to spermatophore transfer.
|
 issued from : K. Hulsemann & A. Fleminger in Mar. Biol., 1990, 105. [p.102, Fig.2, a]. Female: a, genital segment (right lateral view).
|
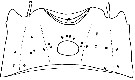 issued from : K. Hulsemann & A. Fleminger in Mar. Biol., 1990, 105. [p.104, Fig.4]. Female: genital segment (dorsal, lateral and ventral patterns separated by dotted line). Integument cut dorsally and laid out flat, showing positions of integumental organs: peg sensilla (o) and pores of integumental glands (filled circle); symbols are larger than organs. Nota: Character divergence in pore signature pattern of the We may reason that a character contributing to a prezygotic reproductive barrier should occur in strongly divergent states among geographically overlapping species. The facts concerning pore signature pattern and geographical relationships match our expectations. Fleminger & Hulsemann (1974) found Pontellina plumata and P. morii to be sympatric across much of the equatorial Indian and Pacific Oceans and P. plumata and P. sobrina to overlap in the eastern equatorial Pacific Ocean. Differences in the pore signature pattern of the two pairs of sympatric species are of the magnitude that distinguishes the two Pontellina species groups. The other geographycally linked species of Pontellina that are allopatric or parapatric in the same region, i.e. P. platychela and P. plumata in the equatorial Atlantic, and P. morii and P. sobrina in the eastern equatorial present weakly differenciated pore signature pattern.
|
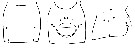 issued from : K. Hulsemann & A. Fleminger in Mar. Biol., 1990, 105. [p.104, Fig.5-7]. Female: genital segment (left to right: dorsal view, right latera viewl and ventral view).Generalized distribution of peg sensilla (o) and pores of integumental glands (filled circle); symbols are larger than organs. Organs shown occurred in at least 40 % of specimens examined (n = 25).
|
 issued from : K. Hulsemann & A. Fleminger in Mar. Biol., 1990, 105. [p.105, Fig.8]. Female: genital segment (left to right: dorsal view, right latera viewl and ventral view) illustrating geographical variation of pore signature pattern; idealized view of integumental organ types, numbers and arrangment. Organs shown are those appearing in at least 40 % of specimens from one regional sample. . Top: dorsal view; middle: right lateral view; bottom: ventral view. Left: types of organs inferred from several intact specimens stained in chlorazole black E and cleared in lactophenol. Symbols: peg sensilla (o); pores of integumental glands (filled circle); symbols are larger than organs. Symbols used for regional samples: (black circle) organs present in 100 % of specimens examined; (clear circle) 80 to 99 %; (black triangle) 60 to 79 %; (white triangle) 40 to 59 %.
|
 issued from : S. Ohtsuka & R. Huys in Hydrobiologia, 2001, 453/454. [p.448, Fig.4, F]. Male (from Japan): modified elements on geniculate A1 of compound segment XXI-XXIII. Scale bar = 0.01 mm
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.48, Figs.1-12]. Female: 1, habitus (dorsal); 2, Mxp; 3, P5; 8, P1; 9, A2. Female immature: 4, habitus (dorsal); 5, P5; 6, P4; 7, Mxp. Male: 10, habitus (dorsal); 11, P5; 12, right A1.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.47, Figs.8-11]. Male: 8, 11, habitus (dorsal and lateral, respectively); 9, A2; 10, P5
|
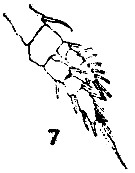 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.47, Fig.7]. Male: 7, P1.
|
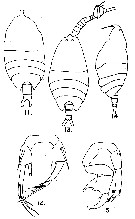 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.238, Pl.34, Figs.11-15]. Female (from equatorial Pacific): 11, habitus (dorsal); 12, P5. Right caudal ramus fused to the anal segment. Male: 13, habitus (dorsal and lateral, respectively); 15, P5. Nota: Posterior lateral margins of the last thoracic segment rounded.
|
 issued from : Mulyadi in Treubia, 2002, 32. [p.158, Fig.59]. Female (from Indonesian waters): a, habitus (dorsal); b, metasomal somite 5 and urosome (dorsal); c, P5; d, variation observed in lateral margin of right caudal ramus (dorsal view). Male: e, P5; f, habitus (dorsal). The present female specimens have various shape of endopd similar to those reported from Pacific Ocean.
|
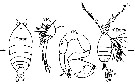 issued from : G. Harding in Key to the adullt pelagic calanoid copepods found over the continental shelf of the Canadian Atlantic coast. Bedford Inst. Oceanogr., Dartmouth, Nova Scotia, 2004. [p.16]. Female & Male. Nota: Female P1 3-segmented. male P5 uniramous.
|
 issued from : N. Phukham in Species diversity of calanoid copepods in Thai waters, Andaman Sea (Master of Science, Univ. Bangkok). 2008. [p.185, Fig.59]. Female (from W Malay Peninsula): a, habitus (dorsal); b, urosome (dorsal); c, P5. Male: d-e, habitus (dorsal and lateral respectively); f, P5. Body length after the drawings: F = 1.440 mm; M = 1.444 mm.
|
 Issued from : J.M. Bradford-Grieve, E.L. Markhaseva, C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, edit. D. Boltovskoy. 1999, Vol. 2, Copepoda; [p.1072, Fig. 7.399: Pontellina plumata]. r = right leg; l = left leg. Female characters (from key, p.960): - Posterior prosome corners extend more than 1/2 way along genital segmen; P5 basis seta extends well short of base of 1st outer edge spine on terminal segment. Male characters (from key, p.960): - Posterolateral corners of prosome, in lateral view, angular and bearing denticle; right chela exopodal segment1-2 extending distally as relatively slender, digitiform process opposing apex of narrow, curved exopodal segment 3.
|
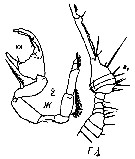 Issued from : M.C. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. After Brodsky, 1962. Male: 1, right A1 (part.); 2, P5.
| | | | | Ref. compl.: | | | Cleve, 1904 a (p.195); Massuti Alzamora, 1942 (p.98, Rem.); Oliveira, 1945 (p.191); Sewell, 1948 (p.392); C.B. Wilson, 1950 (p.303); Hoenigman, 1955 (p.49); Yamazi, 1958 (p.152, Rem.); Demir, 1959 (p.176); Fagetti, 1962 (p.36); Cervigon, 1962 (p.181, tables: abundance distribution); Grice & Hart, 1962, (p.287, 295: Rem.); Duran, 1963 (p.24); Giron-Reguer, 1963 (p.54); V.N. Greze, 1963 a (tabl.2); Sherman, 1963 (p.216: fig.2, p.220: fig.8, p.223: fig.13, p.224: fig.14); Gaudy, 1963 (p.29, Rem.); Björnberg, 1963 (p.61, Rem.); De Decker, 1964 (p.16, 24, 28); De Decker & Mombeck, 1964 (p.13); Heinrich, 1964 (p.89: fig.6, 91: fig.7); Grice & Hulsemann, 1965 (p.225); Mazza, 1966 (p.72); Pavlova, 1966 (p.44); Fleminger, 1967 a (tabl.1); Evans, 1968 (p.14); Delalo, 1968 (p.138); Vinogradov, 1968 (1970) (p.78, 80); Omori, 1969 (p.5, Table 1); Morris, 1970 (p.2301); Park, 1970 (p.478); Timonin, 1971 (p.281, trophic group); Deevey, 1971 (p.224); Salah, 1971 (p.320); Brodsky, 1972 (p.256); Roe, 1972 (p.277, tabl.1, tabl.2); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night, Table II); Apostolopoulou, 1972 (p.328, 370); Heinrich, 1973 (p.95, fig.2); Björnberg, 1973 (p.352, 388); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Weikert, 1975 (p.140, chart); Patel, 1975 (p.660); Lakkis, 1976 (p.84: Rem.); Tranter, 1977 (p.596); Deevey & Brooks, 1977 (p.156, tab.2, Station "S"); Madhu Pratap & al., 1977 (p.138, Table 3: abundance vs. stations); Carter, 1977 (1978) (p.36); Grice & Gibson, 1978 (p.23, tab.8); Dessier, 1979 (p.207); Turner & al., 1979 (p.289, 292, Tab.3, Rem.); Turner & Collard, 1980 (p.527, Tab.1); Svetlichnyi, 1980 (p.28, Table 1, passive submersion); Chen Q-c, 1980 (p.795); Vaissière & Séguin, 1980 (p.23, tab.1); Sreekumaran Nair & al., 1981 (p.493): Madhupratap & al., 1981 (p.266, fig.1e: abundance vs. geographic transect); Vives, 1982 (p.295); Kovalev & Shmeleva, 1982 (p.85); Dessier, 1983 (p.89, Tableau 1, Rem., %); Guangshan & Honglin, 1984 (p.118, tab.); Tremblay & Anderson, 1984 (p.5); De Decker, 1984 (p.317, 361: carte); Scotto di Carlo & al., 1984 (1045); Cummings, 1984 (p.163, Table 2); Regner, 1985 (p.11, Rem.: p.38); Almeida Prado Por, 1985 (p.250); Brenning, 1985 a (p.28, Table 2); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Madhupratap & Haridas, 1986 (p.105, tab.1); M. Lefèvre, 1986 (p.33); Renon, 1987 (tab.2); Brenning, 1987 (p.30, Rem.); Dessier, 1988 (tabl.1); Jimenez-Perez & Lara-Lara, 1988; Lozano Soldevilla & al., 1988 (p.60); Hernandez-Trujillo, 1989 (tab.1); 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Echelman & Fishelson, 1990 a (tab.2); Othman & al., 1990 (p.565, Rem.); Suarez & al., 1990 (tab.2); Hirakawa & al., 1990 (tab.3); Mizushima, 1990 (fig.2, 3, tab.2); Madhupratap & Haridas, 1990 (p.305, fig.4: vertical distribution night/day; fig.7: cluster); Yoo, 1991 (tab.1); Baessa De Aguiar, 1991 (1993) (p.99, 107); Suarez & Gasca, 1991 (tab.2); Suarez, 1992 (App.1); Seguin & al., 1993 (p.23); Hernandez-Trujillo, 1994 (tab.1); Shih & Young, 1995 (p.72); Webber & al., 1996 (tab.1); Suarez-Morales & Gasca, 1997 (p.1525); Go & al., 1997 (tab.1); Park & Choi, 1997 (Appendix); Hure & Krsinic, 1998 (p.103); Padmavati & al., 1998 (p.349); Alvarez-Cadena & al., 1998 (tab.2,4); Hsieh & Chiu, 1998 (tab.2); Noda & al., 1998 (p.55, Table 3, occurrence); Mauchline, 1998 (tab.8, 64); Suarez-Morales & Gasca, 1998 a (p.110); Neumann-Leitao & al., 1999 (p.153, tab.2); Siokou-Frangou, 1999 (p.478); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); Fernandez-Alamo & al., 2000 (p.1139, Appendix); Suarez-Morales & al., 2000 (p.751, tab.1); Lopez-Salgado & al., 2000 (tab.1); Madhupratap & al., 2001 (p. 1345, vertical distribution vs. O2, figs.4, 5: clusters, p.1353); Lo & al., 2001 (1139, tab.I); Dalal & Goswami, 2001 (p.22, fig.2); Hidalgo & Escribano, 2001 (p.159, tab.2); Hidalgo & Escribano, 2001 (p.158, fig.5); Beaugrand & al., 2002 (p.179, figs.5, 6); Hwang & al., 2003 (p.193, tab.2); Vukanic, 2003 (p.139, tab.1); Alvarez-Silva & al., 2003 (p.737: tab.2); Osore & al., 2003 (p.69); Hsiao & al., 2004 (p.326, tab.1); Rezai & al., 2004 (p.489, tab.2); Lan & al., 2004 (p.332, tab.1); Lo & al.*, 2004 (p.218, fig.6); Gallienne & al., 2004 (p.5, tab.3); CPR, 2004 (p.61, fig.181); Lo & al., 2004 (p.89, tab.1); Alvarez-Silva & al., 2005 (p.39); Zuo & al., 2006 (p.163: tab.1); Isari & al., 2006 (p.241, tab.II); Lavaniegos & Jiménez-Pérez, 2006 (p.124: Tab.2, 4, Rem.); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño); Hwang & al., 2006 (p.943, tabl. I); Rakhesh & al., 2006 (p.93, Table 2, spatial distribution); Hwang & al., 2007 (p.25): Cornils & al., 2007 (p.278, Table 2); Dur & al., 2007 (p.197, Table IV); Tseng L.-C. & al., 2008 (p.153, Table 2, occurrence vs geographic distribution); Tseng L.-C. & al., 2008 (p.46, table 2, abundance vs moonsons); Jitlang & al., 2008 (p.65, Table 1); Rakhesh & al., 2008 (p.154, Table 5: abundance vs stations); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.521); Lopez Ibarra, 2008 (p.1, Table 1, 2: Rem.); Fernandes, 2008 (p.465, Tabl.2); C.-Y. Lee & al., 2009 (p.151, Tab.2); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Hidalgo & al., 2010 (p.2089, Table 2); Mazzocchi & Di Capua, 2010 (p.427); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); W.-B. Chang & al., 2010 (p.735, Table 2, abundance); Hsiao S.H. & al., 2011 (p.475, Appendix I); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Guo & al., 2011 (p.567, table 2, indicator); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Pillai H.U.K. & al., 2011 (p.239, Table 3, vertical distribution); Mazzocchi & al., 2011 (p.1163, fig.6, long-term time-series 1984-2006); Uysal & Shmeleva, 2012 (p.909, Table I); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Tseng & al., 2012 (p.621, Table 3: abundance); Lavaniegos & al., 2012 (p. 11, Appendix); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Tseng & al., 2013 (p.507, seasonal abundance); Jagadeesan & al., 2013 (p.27, Table 3, seasonal abundance); Hirai & al., 2013 (p.1, Table I, molecular marker); Lidvanov & al., 2013 (p.290, Table 2, % composition); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Beltran Castro, 2014 (p.46, Table 1, 2: molecular CO1); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Lopez-Ibarra & al., 2014 (p.453, Table 2, biogeographical affinity); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence); El Arraj & al., 2017 (p.272, table 2); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Belmonte, 2018 (p.273, Table I: Italian zones); Abo-Taleb & Gharib, 2018 (p.139, p.149: occurrence); Palomares-Garcia & al., 2018 (p.178, Table 1: occurrence) | | | | NZ: | 18 | | |
|
Carte de distribution de Pontellina plumata par zones géographiques
|
| | | | | | | | | | | | 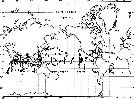 issued from : A. Fleminger & K. Hulsemann in Fishery bulletin, 1974, 72 (1). [p.76, Fig.5]. issued from : A. Fleminger & K. Hulsemann in Fishery bulletin, 1974, 72 (1). [p.76, Fig.5].
Geographical distribution of captures recorded during the present study. |
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.100, Table 46]. issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.100, Table 46].
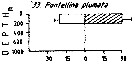
Vertical distribution of Pontellina plumata at the ''40-Mile station'' in the Florida Current (± 25°35'N, 79°27'W).
SL 53: 18 V 1958; SL 55: 21 VII 1958. A: during midday; B;: during midnight. |
 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.311, Fig.4]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.311, Fig.4].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 33- Pontellina plumata.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
| | | | Loc: | | | South Africa (E & W), off Angola, Congo, off S St. Helena Is., off S Ascension Is., G. of Guinea, off E St. Paul Is., Morocco-Mauritania, N Argentina, Brazil, off Rio de Janeiro, Campos Basin, off Natal, off Amazon, Dakar, off Morocco-Mauritania, Canary Is., Azores, Barbados Is., Venezuela, Cariaco Basin, Caribbean Colombia, Caribbean Sea, Yucatan, Jamaica, Yucatan (Ascension Bay), G. of Mexico, Cuba, Florida, Sargasso Sea, off Bermuda: Station S (32°10N, 64°30W), Woods Hole, off Cape Cod, off S Nova Scotia, S Ireland (rare), Ibero-moroccan Bay, Gibraltar, Medit. (Alboran Sea, NW Basin, G. of Lion, Marseille, Ligurian Sea, Tyrrhenian Sea, Napoli, Messina, Malta, Adriatic Sea, Mljet Is., Ionian Sea, Aegean Sea, Lebanon Basin, Alexandria), G. of Suez, G. of Aqaba, Hurghada, Red Sea, Arabian Sea, Arabian Gulf, Lakshadweep Is., Kavaratti Is., Maldive Is., Sri Lanka, Natal, Mascarene Basin, Madagascar (Nosy Bé), Rodrigues Is. - Seychelles, India (Saurashtra coast, W, Gulf of Mannar, Palk Bay), E India (cosatal, Kakinada Bay), Bay of Bengal, Burma, Andaman Sea (Barren Island), W Malay Peninsula (Andaman Sea), Straits of Malacca, G. of Thailand, W Australia, Indonesia-Malaysia (java: off Surabaya, NE Celebes: Manado Bay), Philippines, Gulf of Tonkin, Hong Kong, China Seas (Yellow Sea, East China Sea, Taiwan Strait, South China Sea), Taiwan (S, E, SW, NW, N: Mienhua Canyon, NE), Okinawa, S Korea, Korea Strait, Japan Sea, Japan, Kuchinoerabu Is., Tanabe Bay, off SE Japan, Bikini, off N Carolines Is., Australia (Shark Bay, G. of Carpentaria, Great Barrier, New South Wales), W Pacif. (equatorial), New Caledonia, off New Zealand (N & NE), Moorea Is., Pacif. (N central subtropical), California, W Baja California, Bahia Magdalena, Gulf of California, Bahia de La Paz, Bahia de los Angeles, Islas Marias, G. of Tehuantepec, Zihuatanejo Bay, W Mexico, Clipperton Is., Central America, W Costa Rica, Bahia Cupica (Colombia), off Ecuador, Peru, Chile (N-S, Mejillones Peninsula)
Type locality: 05°N, 21°E. | | | | N: | 292 | | | | Lg.: | | | (16) F: 1,93-1,5; M: 1,73-1,45; (34) F: 1,72-1,68; M: 1,5-1,45; (45) F: 1,75-1,6; M: 1,65-1,5; (46) F: 1,75-1,7; M: 1,62-1,55; (72) F: 1,94; (73) M: 1,6; (91) F: 1,75-1,6; M: 1,65-1,5; (101) F: 1,84-1,43; M: 1,59-1,49; (104) F: 1,89; (151) M: 1,6; (180) F: 1,87-1,79; M: 1,68-1,6; (187) F: 1,94; (199) F: 1,67; (237) F: 1,5-1,9; M: 1,8; (244) F: 1,7; (256) F: 1,6-1,49; M: 1,6-1,26; (275) F: 1,94-1,44; M: 1,92-1,34; (290) F: 1,7-1,85; M: 1,45-1,55; (332) F: 1,7; (334) F: 1,9-1,6; M: 1,65-1,5; (335) F: 1,78-1,64; M: 1,63-1,49; (336) F: 1,78-1,67; M: 1,67-1,5; (530) F: 1,5; M: 1,3; (785) F: 1,58-1,03; (786) F: 1,85-1,75; M: 1,58-1,43; (807) F: 1,56; ? 1,62; (?) F: 1,7-1,8; (991) F: 1,43-1,94; M: 1,34-1,92; (1087) F: 1,45-1,55; M: 1,25-1,30; {F: 1,03-1,94; M: 1,25-1,92} | | | | Rem.: | épipélagique.
Cette espèce présente un cycle nycthéméral inverse (Roehr & Moore, 1965).
Certaines identifications sont considérées comme douteuses (cf. in Fleminger & Hulsemann, 1974, p.75).
Voir aussi les remarques en anglais | | | Dernière mise à jour : 24/10/2022 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 12 juin 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
