|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Calanoidea ( Superfamille ) |
|
|
|
Calanidae ( Famille ) |
|
|
|
Undinula ( Genre ) |
|
|
| |
Undinula vulgaris (Dana, 1849) (F,M) | |
| | | | | | | Syn.: | Undina vulgaris Dana, 1849; 1852; Brady, 1883 (p.53, figs.F,M); T. Scott, 1894 b (p.44); King & Hida, 1955 (p.11);
Calanus vulgaris : Giesbrecht, 1888; 1892 (p.92, 129, 770, figs.F,M); Giesbrecht & Schmeil, 1898 (p.17, Rem. F,M); Thompson & Scott, 1903 (p.232, 242); Cleve, 1904 a (p.186); Carl, 1907 (p.16); Willey, 1919 (p.185); Farran, 1929 (p.207, 216); Mori, 1929 (p.168, figs.F); Dakin & Colefax, 1933 (p.204); Mori, 1937 (1964) (p.19, figs.F,M); Chiba & al., 1957 (p.306); 1957 a (p.11); Delalo, 1968 (p.137); Maiphae & Sa-ardrit, 2011 (p.641, Table 2);
Calanus vulgaris plumosus Wolfenden, 1905 (1906) (p.994, figs.F);
Calanus orientalis Marukawa, 1908;
Stephos perplexus C.B. Wilson, 1950 (p.341, figs.M);
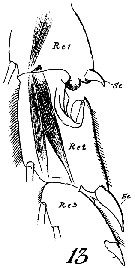
Undinula vulgaris var. taiwanicus : J.-S. Hwang & J.T. Turner, 1995 (p.207, Table 1, 2, fig. 1, 2, 3: swimming behaviour); | | | | Ref.: | | | A. Scott, 1909 (p.16); Sewell, 1912 (p.353, 356: Rem.); 1914 a (p.196, figs.F, Rem.: 3 var.); Früchtl, 1924 b (p.33); Sars, 1925 (p.10); Rose, 1929 (p.9); Sewell, 1929 (p.31, figs.F, juv., Rem.: 2 forms); Wilson, 1932 a (p.29, figs.F,M); Farran, 1936 a (p.77); Dakin & Colefax, 1940 (p.89, figs.F,M); Lysholm & al., 1945 (p.7); Vervoort, 1946 (p.72, fig.F, Rem.); Sewell, 1947 (p.20); C.B. Wilson, 1950 (p.350, fig.M); Carvalho, 1952 a (p.138); Tanaka, 1956 (p.265); 1960 (p.20, Rem.); Fish, 1962 (p.6); Brodsky, 1962 c (p.104, figs.F,M); Cervigon, 1962 (p.181, tables: abundance distribution); Grice, 1962 (p.178, Rem.); Vervoort, 1963 b (p.83, Rem.); Paiva, 1963 (p.14, figs.F); Gaudy, 1963 (p.19, Rem.); Björnberg, 1963 (p.15, figs.F, Juv., Rem.); Kasturirangan, 1963 (p.13, figs.F,M); Chen & Zhang, 1965 (p.31, figs.F,M); Saraswathy, 1966 (1967) (p.75); Owre & Foyo, 1967 (p.32, figs.F,M); Vinogradov, 1968 (1970) (p.78); Koga, 1968 (p.17, fig.: egg); Tanaka, 1969 (p.255, Rem.); Shih & al., 1971 (p.37); Ramirez, 1971 (p.83, figs.F,M); Brodsky, 1972 (1975) (p.73, 90, 120, fig.M); Grice, 1972 (p.178, figs.F,M); Kos, 1972 (Vol.I, figs.F,M, Rem.); Marques, 1973 (p.234); Heinrich, 1973 (p.95); Bradford & Jillett, 1974 (p.6, figs.F,M); Marques, 1974 (p.11); 1976 (p.986); Greenwood, 1976 (p.8, fg.F); Björnberg & al., 1981 (p.618, figs.F,M); Fleminger, 1985 (p.276, 285, Table 1, 4, fig.M, Rem.: A1); 1986 (p.92, 94, 95, figs.F, Rem.: spéciation); Baessa de Aguiar, 1987 (p.39, figs.F,M); Bradford, 1988 (p.74, 76, Rem.); Nishida, 1989 (p.173, table 2, no M dorsal hump); Blades-Eckelbarger, 1991 (p.250, figs.M,F); He & al., 1992 (p.250); Renon, 1993 (p.239); Kim & al., 1993 (p.270); Bradford-Grieve, 1994 (p.44, figs.F, M, fig. 98); Chihara & Murano, 1997 (p.739, Pl.64: F,M); Bradford-Grieve & al., 1999 (p.877, 907, figs.F,M); Bucklin & al., 2003 (p.335, tab.2, fig. 4, Biomol.); Conway & al., 2003 (p.151, figs.F,M, Rem.); G. Harding, 2004 (p.5, 33 figs.F, M); Boxshall & Halsey, 2004 (p.81: fig.F); Mulyadi, 2004 (p.8, 17, figs.F,M, Rem.); Conway, 2006 (p.9: copepodite stages 1-6, Rem.); Vives & Shmeleva, 2007 (p.905, figs.F,M, Rem.); Phukham, 2008 (p.114, figs.F,M); Lacuna & al., 2013 (p.70, Figs.F,M, Rem.); Fornshell & Ferrari, 2014 (p.101, fig.1: Von Vaupel Klein"s organ) | 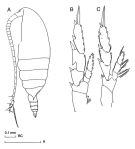 issued from : J.M. Bradford-Grieve in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). New Zealand Oceanographic Institute Memoir, 102, 1994. [p.45, Fig.18]; Female: A, habitus (lateral left side); B, P2; C, P5. Nota: Only one specimen of the typical form was taken from N Tasman Sea. Nota: Posterior corners of prosome pointed. Exopodite 2 of P2 with outer proximal edge invaginate.
|
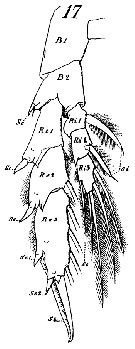
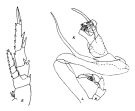 issued from : J.M. Bradford & J.B. Jillett in Crustaceana, 1974, 27 (1). [p.14, Fig.2,E,K]. Female: E, P2. Male: K, P5 (L = left, R = right).
|
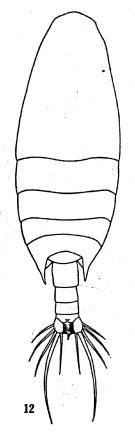 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.3, 12]. Female (from E China Sea): 12, habitus (dorsal).
|
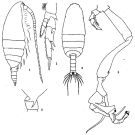 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.4, 1-5]. Female: 1, habitus (lateral right side); 2, postero-lateral angles of last thoracic segment (lateral left side); 3, P2 (anterior). Male: 4, habitus (dorsal); 5, P5 (anterior).
|
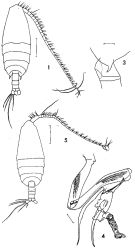 issued from : F.C. Ramirez in Revta Mus. La Plata, Seccion Zool., 1971, XI. [Lam.I, Figs.1,3, 4, 5]. Female (from off Mar del Plata): 1, habitus (dorsal); 3, posterior part cephalothorax and genital segment (lateral left side). Male: 4, P5; 5, habitus (dorsal). Scale bars in mm: 0.5 (1, 5); 0.1 (3, 4);
|
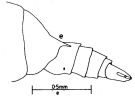 issued from : J.G. Greenwood in Proc. R. Soc. Qd, 1976, 87. [p.9, Fig.3, e]. As Undinula vulgaris forma typica. Female (from Moreton Bay, E Australia): e, posterior metasomal segment and urosome (lateral left side).
|
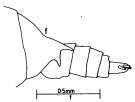 issued from : J.G. Greenwood in Proc. R. Soc. Qd, 1976, 87. [p.9, Fig.3, f]. As Undinula vulgaris forma gisebrechti. Female (from Moreton Bay, E Australia): posterior metasomal segment and urosome (lateral left side). Nota: Variety in which last metasomal segment of female with spine on left side is doubled by addition of a ventrally directed spine. Both varieties were represented in Moreton Bay specimens. There was no evidence of two size classes as noted in other regions (see Sewell, 1929; Tanaka, 1960).
|
 Issued from : R.B.S. Sewell in Spolia Zeylanica, 1914, 9. [Pl.XVII, Figs.1-2. Female (from Gulf of Mannar): 1, left posterior thoracic margin (lateral view); 2, idem (dorsal view).]
|
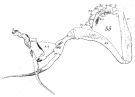 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.8, Fig.35]. As Calanus vulgaris. Male: 35, P5 (anterior surface).
|
 issued from : R.B.S. Sewell in Mem. Indian Mus., 1929, X. [p.37, Fig.8]. Female (from Nicobar Is.): a, posterior thoracic margin and urosome , lateral left side (forma minor); b, idem (forma major); c, P2 (forma minor); d, idem (forma major); e, P5 abnormal (hermaphrodite form).
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.6, Figs.1-3]. As Calanus vulgaris. Female: 2, P5; 3, habitus (lateral). Male: 1, habitus (lateral).
|
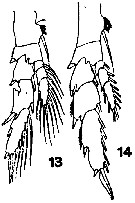 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.5, Figs.13-14]. As Calanus vulgaris. Female: 13, P2; 14, P3.
|
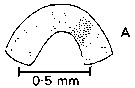 issued from : J. Mauchline in J. mar. biol. Ass, U.K., 1977, 57 (4). [p.980, Fig.4, A]. A, Integumental organs on fused 4th and 5th thoracic segments.
|
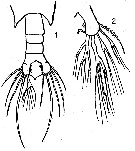 issued from : R. Gaudy in Rec.Trav. Stn mar. Endoume, 1963, 45, Bull. 30 (45). [p.40, Pl.I,, Figs.1-2]. Female (from N-S Brazil): 1, urosme (with anomalies of caudal ramus seta); 2, caudal ramus with anomalies of setae.
|
 issued from T. Mori in Zool. Mag. Tokyo, 1929, 41 (486-487). [Pl. III, Figs. 16-19, 24]. As Calanus vulgaris. Female (from Chosen Strait, Korea-Japan): 16, last thoracic and 1st-2nd abdominal segments; 17, P2; 18, P3; 19, last thoracic and 1st abdominal segment; 24, P5.
|
 issued from T. Mori in Zool. Mag. Tokyo, 1929, 41 (486-487). [Pl. IV, Fig.2]. As Calanus vulgaris. Male: 2, P5.
|
 issued from : A. Fleminger in Mar. Biol., 1985, 88. [p.284, Fig.6 A]. Male (from Atlantic tropical): Left A1 proximal segments (ventral view); Nota: see remarks in Calanus s.l. pacificus californicus (Fleminger, 1985, p.275) concerning the dimorphism in the female A1.
|
 Issued from : G.S. Brady in Rep. Scient. Results Voy. Challenger, Zool., 1883, 8 (23). [Pl.XVIII, Fig.6]. As Undina vulgaris. Female: 6, habitus (lateral).
|
 Issued from : G.S. Brady in Rep. Scient. Results Voy. Challenger, Zool., 1883, 8 (23). [Pl.XV, Figs.12, 13]. As Undina vulgaris. Female: 12, A1; 13, P2.
|
 Issued from : G.S. Brady in Rep. Scient. Results Voy. Challenger, Zool., 1883, 8 (23). [Pl.XV, Figs.11, 14, 15]. As Undina vulgaris. Male: 11, A1; 14, P5; 15, urosome.
|
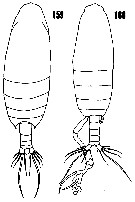 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1967, 1, Part 1: Copepoda. [p.33, Figs.159-160]. Female (from Florida Current): 159: habitus (dorsal). Male: 160, habitus (dorsal).
|
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1967, 1, Part 1: Copepoda. [p.33, Fig.165]. Female: 165, P5.
|
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.16, Fig.43]. Male: 43, P5.
|
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.17, Fig.46]. Female: 46, P2.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.7, Fig.2]. As Calanus vulgaris. Female: 2, A2 (postrior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 8, Fig.13]. Female: 13, exopodite 1-3 of P2 (posterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 8, Fig.17]. As Calanus vulgaris. Female: 17, P5 (anterior view).
|
 issued from : N. Phukham in Species diversity of calanoid copepods in Thai waters, Andaman Sea (Master of Science, Univ. Bangkok). 2008. [p.197, Fig.71]. Female (from W Malay Peninsula): a, habitus (dorsal); b-c, urosome (dorsal and lateral, respectively); e, P2. Copepodite. Male: f, habitus (dorsal); g, same with P5; h, P5. Body length after the drawings: F = 2.667 mm; M (1! = 2.482 mm; M (2) = 2.293 mm.
|
 issued from : J.M. Bradford-Grieve, E.L. Markhaseva & C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, Edit. D. Boltovskoy. 1999. Vol.2. Copepoda. [p.982, Fig. 7.123]. Male: habitus (lateral); P5 (L = left leg; r = right leg). Nota: Left P5 highly modified, of complex structure (prehensile). Coxa of P5 with inner edge without teeth. Exopodite 2 of P2 with invaginated outer proximal border.
|
 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.18, Fig.6]. Female (from Indonesian Seas): a, habitus (dorsal); b-f, P1 to P5, respectively. Male: g, habitus (dorsal); h, P5.
|
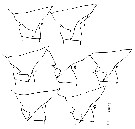 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.19, Fig.7]. Female: a-g, variations of lert posterior ends of last thoracic segment. Nota: The subspecies typica is identified by the asymmetrical posterolateral ends of corners as in fig.a, b. The subspecies giesbrechti characterizes by the double spines on the left posterolateral end of posterior thoracic segment, the upper spine projects straightly backwards, while the lower curved downwards; right side as in typica, single and curved downwards (fig.f). The subspecies zeylonica bears double spines on the left posterolateral end as in giesbrechti, but the whole thoracic corner considerably thickened; the spine on the right side is straight and points backwards.
|
 Issued from : J.H. Wickstead in Parasitology, 1963, 53. [p.294, Fig.1]. Elliobiopsis chattoni on Undinula vulgaris var. major (from Zanzibar Channel).A: Female with six parasites attached. B: young parasite on A1. length of copepod: 2.78 mm; In B parasite measures 8.3 x 10.9 µ. Nota: Total individuals parasited: 428 females and 337 males. After the author it was found that the parasite affected egg production in the ovaries. The suggestion is that it produces a starvation effect on the copepod, this effect being reversible.
|
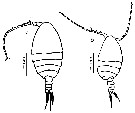 ssued from : M.L.D.G. Lacuna, D.C. Sagrado, R.O. Mejorada, D.D. Simyunn & M.J.J. Pueblos in ABAH Bioflux, 2013, 5 (1). [p.71, Fig.23]. Female (from Butuuan Bay, Mindanao): a, habitus (dorsal). Male: b, habitus (dorsal). Nota: Both sexes are semi-transperent in coloration in leve specimens. The posterior margins of the metasome in female are drawn out into a spine
|
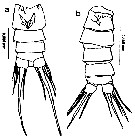 ssued from : M.L.D.G. Lacuna, D.C. Sagrado, R.O. Mejorada, D.D. Simyunn & M.J.J. Pueblos in ABAH Bioflux, 2013, 5 (1). [p.71, Fig.24]. Female: a, urosome (ventral). Male: b, urosome (dorsal). Nota: Caudal rami of females wider comparated to the male, each ramus with 5 setae, 2nd seta the longest, 5th seta shortest among the other setae.
|
 ssued from : M.L.D.G. Lacuna, D.C. Sagrado, R.O. Mejorada, D.D. Simyunn & M.J.J. Pueblos in ABAH Bioflux, 2013, 5 (1). [p.72, Figs.25, 26]. Female: a, left A1; b, right A1. Male: c, left A1; d, right A1. Noya: Left A1 female 23-segmented, right A1 with 24 segments. Left A1 male 23-segmented, right A1 where geniculation occurs, much shorter than the left, with 21 segments; the segments associated with the geniculation occurred from 2nd to 6th segments.
|
 ssued from : M.L.D.G. Lacuna, D.C. Sagrado, R.O. Mejorada, D.D. Simyunn & M.J.J. Pueblos in ABAH Bioflux, 2013, 5 (1). [p.73, Fig.27]. Female: a, P1; b, P2 (red arrow indicates a deep notched; c, P3; d, P4; e, P5.
|
 ssued from : M.L.D.G. Lacuna, D.C. Sagrado, R.O. Mejorada, D.D. Simyunn & M.J.J. Pueblos in ABAH Bioflux, 2013, 5 (1). [p.73, Fig.28]. Male: a, P1; b, P2 (red arrow indicates a deep notched); c, P3; d, P4; e, P5. Nota: The right P5 shorter, with a 3-segmented exopod and endopod, exopod much longer than endopod; left P5 uniramous, with 2-segmented exopod
|
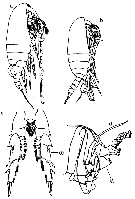 Issued from : M. Chihara & M. Murano in An Illustrated Guide to Marine Plankton in Japan, 1997. [p.743, Pl. 64, fig.82 a-d]. Female: a, habitus (lateral); c, P5. Male: b, habitus (lateral); d, P5. Nota: Numbers show species characteristics.
|
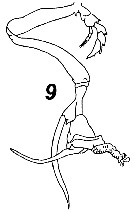 Issued from : V.N. Andronov in Russian Acad. Sci. P.P. Shirshov Inst. Oceanol. Atlantic Branch, Kaliningrad, 2014. [p.73, Fig.19, 9]. Undinula vulgaris after Owre & Foyo, 1967. Male P5.
|
 Issued from : A. Fleminger in UNESCO Techn. papers mar. Sci., 1986, 49. [p.95, Fig.9]. Undinula vulgaris s.l., adult female. Scatter diagram showing length of the right fifth pediger-bearing thoracic segment (ThV) plotted against standard body length (prosome) in six geographical populations. Open suares and crosses represent subspecies zeylonica from Pago Pago and the Marshall Islands, respectively. Filled triangles, filled circles and filled squares represent subspecies giesbrechti from Wallacea north and south of equator, respectively. Open circles represent subspecies Typica from Indian Ocean and eastern Pacific. Inverted open triangles represent subspecies Typica from Atlantic Ocean. Small figures on the left are right lateral views of ThV and genital segment; distribution of cuticular glands and openings shown as columnar structures and dots. Shaded area indicates storage of glandular products. Small figures on right are left lateral and dorsal views of ThV and genital segment. From the top down, subspecies zeylonica, giesbrechti, typica; the fourth figure down on the left is subspecies typica from the Atlantic.
|
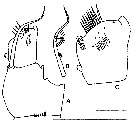 Issued from : J.A. Fornshell & F.D. Ferrari in Crustaceana, 2014, 87 (1). [p.105, Fig.1, A-C]. Von Vaupel Klein's organ on right endopodal segment 1 and right basal seta of P1. A, anterior, distal up; B, right basal seta; C, proximal endopodal segment Scale line; short = 45 µm; long = 65 µm. The Von Vaupel Klein organ is an association (srtucture) of the distal seta from basis and the proximal endopodal segment of P1. This structure shows significant variability among many gymnoplean copepods, in the shape of the distodorsal corner of the proximal endopodal segment, presence and location of denticles on the anterior face of the segment, presence and size of denticles along the distal margin of the segment , number of pores on the segment, shape of the seta that originates on the basis, and the morphology of the basis at the origin of the seta. The function of this complex structure is not known.
|
 Issued from : M.C. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. After Brodsky, 1962. Female: 1-2, habitus (dorsal and lateral, respectively); 3-4, left corners of the thoracic posterior segment; 5, same on he right side; 6, P2; 7, P5. Male: 8, habitus (dorsal); 9, P5 (np = right)
| | | | | Ref. compl.: | | | Wilson, 1942 a (p.210); Oliveira, 1945 (p.191); Sewell, 1948 (p.322, 325, 357, 381, 390, 395, 403, 407, 414, 422, 433, 442, 450, 456, 459, 463, 465, 489, 491); M.W. Johnson, 1949 (238, fig.2); Krishnaswamy, 1953 (p.109); Kott, 1957 (p.5, Rem.: p.8); Yamazi, 1958 (p.147, Rem.); Fagetti, 1962 (p.9); Ganapati & Shanthakumari, 1962 (p.7, 16); Grice & Hart, 1962 (p.287, table 3); Wickstead, 1963 (p.293, var. major, parasite); De Decker, 1964 (p.16, 25, 27); De Decker & Mombeck, 1964 (p.14); Roehr & Moore, 1965 (p.565, vertical distribution); Neto & Paiva, 1966 (p.18, Table III); Fleminger, 1967 a (tabl.1); De Decker, 1968 (p.45); Park, 1970 (p.474); Morris, 1970 (p.2300); Itoh, 1970 (tab.1); Deevey, 1971 (p.225); Timonin, 1971 (p.281, trophic group); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night, Table II: %, Table IV); Subbaraju & Krishnamurphy, 1972 (p.25, 26 ); Binet & al., 1972 (p.73); Brodsky, 1972 (p.256); Björnberg, 1973 (p.283, 389); Ikeda, 1974 (p.1, respiration, ammonia excretion rate); Patel, 1975 (p.659); Peterson, 1975 (p.25, abundance); Ikeda, 1976 (p.51, respiration rate); Sale P.F. & al., 1976 (p.59, table 3: as Undulina); Deevey & Brooks, 1977 (p.256, Table 2, Station "S"); Tranter, 1977 (p.596); Reeve & al., 1977 (p.105, mercury & copper vs. respiration & excretion); Carter, 1977 (1978) (p.35); Stephen & Iyer, 1979 (p.228, tab.1, 3, 4, figs.3, 4); Binet, 1979 (p.400, fig.2); Dessier, 1979 (p.49, 201); Gerber & Gerber, 1979 (p.33, fecal pellets); Chen Q-c, 1980 (p.795); Svetlichnyi, 1980 (p.28, Table 1, passive submersion); Sreekumaran Nair & al., 1981 (p.493); Madhupratap & al., 1981 (p.262, Table 2: abundance vs. thermocline); Madhupratap & al., 1981 (p.266, fig.2d: abundance vs. geographic transect); Kovalev & Shmeleva, 1982 (p.82); Vidal & Whitledge, 1982 (p.77, respiration vs. latitude); Vives, 1982 (p.290); Smith S.L., 1982 (p.1347, Table 5, grazing); Dessier, 1983 (p.89, Tableau 1, Rem., %); Tranter & Dagg, 1983 (p.16, 22); Moore E.A. & Sander, 1983 (p.113, fig.1: variation % sex ratio); Tremblay & Anderson, 1984 (p.4); Cummings, 1984 (p.163, Table 2); De Decker, 1984 (p.315, 366: chart); Guangshan & Honglin, 1984 (p.118, tab.); Stephen, 1984 (p.161, 169, Distribution vs thermocline & geographic); Binet, 1984 (tab.3); 1985 (p.85, tab.3); Brenning, 1985 a (p.23, Table 2); Kinzer & Schulz, 1985 (tab.4); Almeida Prado Por, 1985 (p.250); Fleminger, 1986 (p.84, , Rem. p.92); Brinton & al., 1986 (p.228, Table 1); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Madhupratap & Haridas, 1986 (p.105, tab.1); M. Lefèvre, 1986 (p.33, 40); Turner, 1986 a (p.1, feeding-fecal pellets); Jimenez-Perez & Lara-Lara, 1988; Dessier, 1988 (tabl.1); Williams D. & al., 1988 (p.580); Hernandez-Trujillo, 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Le Borgne & al., 1989 (p.341, respiration, exccretion); Echelman & Fishelson, 1990 a (tab.2); Hirakawa & al., 1990 (tab.3); Madhupratap & Haridas, 1990 (p.305, fig.6: vertical distribution night/day; fig.7: cluster); Schulz, 1990 (p.188); Suarez & al., 1990 (tab.2); Othman & al., 1990 (p.561, 563, Table 1); Lindo, 1991 (tab.3); Dai & al., 1991 (p.47); Samba Diouf, 1991 (p.103, tab.1: Rem.); Hirakawa, 1991 (p.373); Yoo, 1991 (tab.1); McKinnon, 1991 (p.471); Suarez & Gasca, 1991 (tab.2); Yen & al., 1992 (p.495, tab.1, mechanoreception); Huntley & Lopez, 1992 (p.201, Table B1, growth rate, temperature-dependent production); Suarez, 1992 (App.1); C. Park & Landry, 1993 (p.415, egg production); Hirakawa & al., 1995 (tab.2); Shih & Young, 1995 (p.68); Roff & al., 1995 (p.165, Table 1, 3, 5, nauplii development times, growth rates, nauplii vs. bacteriovory); Webber & Roff, 1995 a (p.481, Table.1, 2, 3, 4, 5, 6, 7, 8, biomass; production); Webber & al., 1996 (tab.1); Madhupratap & al., 1996 (p.866); Padmavati & Goswami, 1996 a (p.85, fig.2, 3, vertical distribution); Kotani & al., 1996 (tab.2); Lenz & al., 1996 (p.1, behaviour); Go & al., 1997 (tab.1); Suarez-Morales & Gasca, 1997 (p.1525); Padmavati & al., 1998 (p.349); Hwang & al., 1998 (tab.II); Alvarez-Cadena & al., 1998 (tab.1,2,3,4); Hopcroft & al., 1998 (tab.2); Hsieh & Chiu, 1998 (tab.2); Wong & al, 1998 (tab.2); Noda & al., 1998 (p.55, Table 3, occurrence); Smith S. & al., 1998 (p.2369, Table 6, moonsoon effects); Mauchline, 1998 (tab.18, 25, 26, 27, 29, 30, 45, 47, 64); Suarez-Morales, 1998 (p.345, Table 1); Suarez-Morales & Gasca, 1998 a (p108); Achuthankutty & al., 1998 (p.1, Table 2, seasonal abundance vs monsoon); Stephen, 1998 (p.339, 340, chart); Neumann-Leitao & al., 1999 (p.153, tab.2); Hassett & Boehlert, 1999 (p.571); Hernandez-Trujillo, 1999 (p.284, tab.1); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); Dolganova & al., 1999 (p.13, tab.1); Lopes & al., 1999 (p.215, tab.1); Lapernat, 2000 (tabl.3, 4); Fernandez-Alamo & al., 2000 (p.1139, Appendix); Suarez-Morales & Gasca, 2000 (1247, tab.1); Suarez-Morales & al., 2000 (p.751, tab.1); Lopez-Salgado & al., 2000 (tab.1); Lenz & al., 2000 (p.337); Madhupratap & al., 2001 (p. 1345, vertical distribution vs. O2, figs.4, 5: clusters); Lo & al., 2001 (1139, tab.I); Greenwood & al., 2002 (p.17, Table 2); Dunbar & Webber, 2003 (tab.1); Osore & al., 2003 (p.69); Hwang al., 2003 (p.193, tab.2); Hsiao & al., 2004 (p.325, tab.1); Hsieh & al. 2004 (p.397,tab. 1); Rezai & al., 2004 (p.490, tab.2); Lan & al., 2004 (p.332, tab.1, tab.2); Lo & al.*, 2004 (p.218, tab.1,fig.6); Gallienne & al., 2004 (p.5, tab.3); CPR, 2004 (p.64, fig.193); Lo & al., 2004 (p.89, tab.1); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Kazmi, 2004 (p.230, Rem.: p.232); Dias & Bonecker, 2005 (p.100 + poster); Smith & Madhupratap, 2005 (p.214, tab.4); Prusova & Smith, 2005 (p.76); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, bfig.7, seasonal abundance vs El-Niño, Rem.: p.70); Zuo & al., 2006 (p159, tab.1, 3, fig.9, spatial abundance, fig.8: stations group); Hwang & al., 2006 (p.943, tab.I); ; Zhang W & al., 2006 (p.553, Table 2, grazing rates); Rakhesh & al., 2006 (p.93, Table 2, spatial distribution); Dias & Araujo, 2006 (p.33, Rem., chart); Lavaniegos & Jiménez-Pérez, 2006 (p.158, tab.2, 4, Rem.); Cornils & al., 2007 (p.278, Table 2); Hwang & al., 2007 (p.23); Jitlang & al., 2008 (p.65, Table 1); McKinnon & al., 2008 (p.843: Table.I, p.848: Tab. IV); Rakhesh & al., 2008 (p.154, Table 5, abundance vs stations); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.513, 518); Lopez Ibarra, 2008 (p.1, Table 1, fig.11: abundance); Fernandes, 2008 (p.465, Tabl.2); Waggett & Buskey, 2008 (p.111, Table 1); Lan Y.C. & al., 2008 (p.61, Table 1, % vs stations); Tseng L.-C. & al., 2008 (p.153, Table 2, fig.5, occurrence vs geographic distribution, indicator species); Tseng & al., 2008 (p.402, Table 2); Tseng L.-C. & al., 2008 (p.46, table 2, abundance vs moonsons); Pagano, 2009 (p.116); C.-Y. Lee & al., 2009 (p.151, Tab.2); Miyashita & al., 2009 (p.815, Tabl.II); Tseng & al., 2009 (p.327, fig.5, feeding); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Hwang & al., 2009 (p.49, fig.4, 5); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Cornils & al., 2010 (p.2076, Table 3); Sun & al., 2010 (p.1006, Table 2); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Dias & al., 2010 (p.230, Table 1); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); W.-B. Chang & al., 2010 (p.735, Table 2, abundance); Hsiao & al., 2010 (p.179, Table I, II, III, trace metal concentration); Xu & Gao, 2011 (p.514, figs.3, 4, Table 2: optimal salinity); Guo & al., 2011 (p.567, table 2, indicator); Pillai H.U.K. & al., 2011 (p.239, Table 3, vertical distribution); Tutasi & al., 2011 (p.791, Table 2, abundance distribution vs La Niña event); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Hsiao S.H. & al., 2011 (p.475, Appendix I); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Hsiao & al., 2011 (p.317, Table 2, fig.6, indicator of seasonal change); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Tseng L.-C. & al., 2011 (p.239, Table 3, abundance %); Hsiao & al., 2011 (p.232, Table 1: abundance, %, Table 2: trace metal concentration, vs transect); Beltrao & al., 2011 (p.47, Table 1, density vs time); Pagano & al., 2012 (p.538, Table 1); Saitoh & al., 2011 (p.85, Table 5); Johan & al., 2012 (2013) (p.1, Table 1); Mulyadi & Rumengan, 2012 (p.202, Rem.: p.204); Lavaniegos & al., 2012 (p. 11, Table 1, seasonal abundance, Appendix); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Tseng & al., 2012 (p.621, Table 3: abundance, indicator species); Teuber & al., 2013 (p.28, Table 1, respiration rates); Teuber & al., 2013 (p.1, Table 1, 2, 3, fig.5, abundance vs oxygen minimum zone, respiration rates, enzyme activity); Palomares-Garcia & al., 2013 (p.1009, Table I, fig.7, abundance vs environmental factors); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Hsiao & Fang, 2013 (p.175, Hg bioaccumulation); Tseng & al., 2013 (p.507, seasonal abundance); Rakhesh & al., 2013 (p.7, Table 1, 4, abundance vs stations); Jagadeesan & al., 2013 (p.27, Table 3, 5, 6, fig.11, seasonal variation); Anjusha & al., 2013 (p.40, Table 3, abundance & feeding behavior); Jungbluth & Lenz, 2013 (p.630, Table I); Mendoza Portillo, 2013 (p.37: Fig.7, seasonal dominance); Tseng & al., 2013 a (p.1, Table 3, 5, abundance); Lidvanov & al., 2013 (p.290, Table 2, % composition); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production, Table 4: % vs. season, fig.8); Nakajima & al., 2015 (p.19, Table 3: abundance); Bode & al., 2015 (p.268, Table 1, 2, figs.3, 4, chemical components, trophic level, geographic zone); Rojas-Herrera & al., 2016 (p.40, Table 2: temporal abundance); Kiko & al., 2016 (p.2241, fig.5, Table 3, respiration, ammonium excretion); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence); Ohtsuka & Nishida, 2017 (p.565, Table 22.1); El Arraj & al., 2017 (p.272, table 2, seasonal composition); Belmonte, 2018 (p.273, Table I: Italian zones); Dias & al., 2018 (p.1, Tables 2, 4, 5: vertical distribution, abundance vs. season); Palomares-Garcia & al., 2018 (p.178, fig.3: relative frequency, Table 1: forgetted); Bode & al., 2018 (p.840, Table 1, fig.5, respiration & ingestion rates, depth); Bode & al., 2018 (p.66, fig.5, vertical distribution %, Rem). Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions). | | | | NZ: | 18 | | |
|
Carte de distribution de Undinula vulgaris par zones géographiques
|
| | | | | | | | | | | |  issued from : R. Stephen in Pelagic Biogeography ICoPB II. Proc. 2nd Int. Conf. Final report of SCOR/IOC working group 93. 9-14 July 1995. Workshop Rep. No.142. UNESCO, 1998. issued from : R. Stephen in Pelagic Biogeography ICoPB II. Proc. 2nd Int. Conf. Final report of SCOR/IOC working group 93. 9-14 July 1995. Workshop Rep. No.142. UNESCO, 1998.
P.340, Fig.3: Distribution of Undinula vulgaris in the Indian Ocean. |
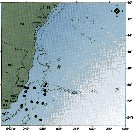 issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.33]. issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.33].
Chart of occurrence in Brazilian waters (sampling between 22°-23° S). |
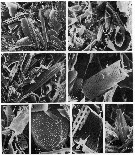 issued from : J.T. Turner in P. S. Z. N. I: Mar. Ecol?, 1986, 7 (1). [p.6, Fig.4]. issued from : J.T. Turner in P. S. Z. N. I: Mar. Ecol?, 1986, 7 (1). [p.6, Fig.4].
Contents of fecal pellets from Undinula vulgaris CV copepodites from Station B-2 (off W Mississipi River) on 11 December 1981.
a: unidentified centric diatom (CD) and Chaetoceros spp spines (Ch); b: Chaetoceros spp. spines (Ch) and fragments of pennate diatoms (PD), Thalassionema nitzschoides (Tn); d, and e: Dactyliosolen sp. (D); f: Prorocentrum sp. cell; g, fragments of Dactyliosolen sp. (D) and Chaetoceros spp. spine (Ch); h: Nitzschia sp. cell (N). |
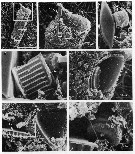 issued from : J.T. Turner in P. S. Z. N. I: Mar. Ecol?, 1986, 7 (1). [p.10, Fig.8]. issued from : J.T. Turner in P. S. Z. N. I: Mar. Ecol?, 1986, 7 (1). [p.10, Fig.8].
Contents of a single fecal pellet from Undinula vulgaris female from station off Mississipi River (3 December 1982).
a: whole pellet (background in glass fiber filter); b: high magnification of area in upper box in a., showing peridinium sp. (P) cell; c: high magnification of area in box at right in b., showing cells of a small centric (SC) and a pennate diatom (PD); d: high magnification of area in box at left in b., showing Thalassionema nitzschoides fragment (Tn); e: high magnification of area in middle box in a., showing Thalassiosira fragment (Th); f: high magnification of area in lower box in a., showing Thalassiosira sp. fragments (Th); g: high magnification of area in box in f., showing a small centric diatom (SC). |
 Issued from : S.-H. Hsiao, J.-S. Hwang & T.-H. Fang in Crustaceana, 2010, 83 (2). [p.184, Table I]. Issued from : S.-H. Hsiao, J.-S. Hwang & T.-H. Fang in Crustaceana, 2010, 83 (2). [p.184, Table I].
Copepods collected from the sea around northern Taiwan and from southern East China Sea extending to the Okinawa Trough, to study possible spatial heterogeneity. The content of the same metal shows considerable variation both intra- and inter-specifically. The metal concentrations in males are higher than in females. Copepod metal quota display spatial variation: coastal water > southern East China Sea > Kuroshio water, suggesting that the metal contents of copepods are influenced by the water quality of their marine environment. |
 Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6]. Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6].
Variations in the most abundant copepod species (mean ± SE) along the transect in the boundary waters between the northern part Taiwan Strait and the East China Sea i March (black bar) and October (grey bar) 2005 (Mann-Whitney U test, sig. ***P <0.001.
See drawings of Hydrological conditions and superficial marine currents in Calanus sinicus. |
 issued from : J. Yen, P.H. Lenz, D.V. Gassie & D.K. Hartline in J. Plankton Res., 1992, 14 (4). [p.502, Fig. 4]. issued from : J. Yen, P.H. Lenz, D.V. Gassie & D.K. Hartline in J. Plankton Res., 1992, 14 (4). [p.502, Fig. 4].
Neural activity from the A1 of Undinula vulgaris from Kaneohe Bay, Hawaii.
Left side: spontaneous background activity. Right side: responses to a variety of mechanical stimuli (tap). All recordings are 40 ms long.
Compare with Labidocera madurae, Euchirella curticauda, Acartia fossae, Pleuromamma xiphias. |
 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 66- Undinula vulgaris.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
 Issued from : C.O. Dias, A.V. Araujo, S.C. Vianna, L.F. Loureiro Fernandes, R. Paranhjos, M.S. Suzuki & S.L.C. Bonecker in J. Mar. Biol. Assoc. U.K., 2015, 95 (3). Issued from : C.O. Dias, A.V. Araujo, S.C. Vianna, L.F. Loureiro Fernandes, R. Paranhjos, M.S. Suzuki & S.L.C. Bonecker in J. Mar. Biol. Assoc. U.K., 2015, 95 (3).
[p.490, Table 2].
In list of copepod from The Campos Basin (20.5 and 24°S off the central Brazilian coast. Abundance, biomass and production od species collected by horizontal subsurface hauls ( 1 m depth) , by a Multinet (200 µm mesh aperture) in dry and and rainy seasons.
For calculations see text p.485. Map (figt.1) see to Clausocalanus furcatus for the sampling stations. |
 Issued from : C.O. Dias, A.V. Araujo, S.C. Vianna, L.F. Loureiro Fernandes, R. Paranhjos, M.S. Suzuki & S.L.C. Bonecker in J. Mar. Biol. Assoc. U.K., 2015, 95 (3). [p.485]. Issued from : C.O. Dias, A.V. Araujo, S.C. Vianna, L.F. Loureiro Fernandes, R. Paranhjos, M.S. Suzuki & S.L.C. Bonecker in J. Mar. Biol. Assoc. U.K., 2015, 95 (3). [p.485].
Calculation of biomass and daily copepod production.
See Table 2 for all species studied. |
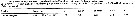 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1].
Length-weight regression of Scolecithrix danae in oceanic waters of Discovery Bay (Jamaica) in 60- and 200 m vertical hauls, using 200- and 64-µm mesh aperture of plankton nets, from June 1989 to July 1991 (monthly samples). |
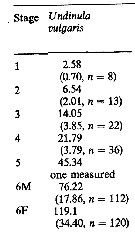 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 2]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 2].
Mean biomass (µg) of copepodite stages for Undinula vulgaris from Discovery Bay (Jamaica), with standard deviation and n number of individuals measured. |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 3]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 3].
Stage weight increments (In mmax mmin-1) for Undinula vulgaris from Discovery Bay (Jamaica), calculated from mean biomass of each stage. |
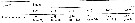 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 4]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 4].
Stage duration (days) for copepodite stages of Undinula vulgaris from Discovery Bay (Jamaica). n: number of observations. |
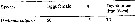 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 5]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 5].
Mean number of eggs per female for Undinula vulgaris from Discovery Bay (Jamaica) and corrected egg development time in days. n number of observations. |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 6]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 6].
Estimates of generation (in days) from egg to first egg clutch for Undinula vulgaris from Discovery Bay (Jamaica) based on minimum and maximum development time of individual life history stages. Centering indicate Max and Min values identical. |
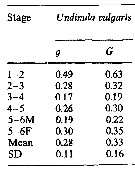 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1].
Growth coefficient (g) and daily specific growth rate (G) for Undinula vulgaris from Discovery Bay (Jamaica). |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1].
Undinula vulgaris from Discovery Bay (Jamaica).
AFDW: ash-free dry weight. |
 Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.170, Table 5]. Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.170, Table 5].
Summary observations on naupliar bacterivory on FLB (fluorescently labelled bacteria) a partir Escherichia coli (±0.7 µm3 cell volume) in Discovery Bay (Jamaica).
Fluorescence levels: 0 = no materal for observation; - = no fluorescence observed; + = low fluorescence due to few FLB observe; * = highly fluorescence due to many FLB. |
 Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, in Table 1). Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, in Table 1).
Cf. explanations of these measures in Calanoides natalis from the same authors. |
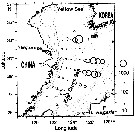 Issued from : T. Zuo, R. Wang, Ya-qu. Chen, Sh-wu Gao & Ke. Wang in J. Mar. Syst., 2006, 59. [p.170, Fig.9F]. Undinula vulgaris Issued from : T. Zuo, R. Wang, Ya-qu. Chen, Sh-wu Gao & Ke. Wang in J. Mar. Syst., 2006, 59. [p.170, Fig.9F]. Undinula vulgaris
Spatial of representative species abundance with isobaths shown in gray.
The circles are proportional in diameter to square root value of abundance.
Copepods collected vertically with plankton net (200 µm mesh aperture) from a depth near the bottom to the surface in autumn 2000 (18 October to 21 November).
Body length (mm): 2.88. |
 Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.209: Table 1, p.210: Table 2]. As Undinula vulgaris var. taiwanicus. Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.209: Table 1, p.210: Table 2]. As Undinula vulgaris var. taiwanicus.
Table 1: Experimental protocol.
Table 2: Swimming versus non-swimming behaviour, and behavioural transitions per minute. Means (bold type), ranges, and standard deviations (SD). Number of repeated observations = 500.
Copepods collected in surface net from waters near Pitou (NE coast Taiwan: 25°N, 122°E) in daylight in July 1993. By cinematographic techniques (see references in Costello & al., 1990; Huang & al., 1993; Turner & al., 1993).
Compare with Oithona nana, O. similis, Oncaea venusta, Macrosetella gracilis, Temora turbinata by the same authors.
Rem: This species frequently alternated between upward motion and sinking at rest, making feeding current about a fourth of the time. There was considerable intraspecific variability between individual animal (it was impossible to statistically test the significance of the variation).
the species alternated between short periods of swimming and non-swimming, spending on average, almost a quarter of its time swimming, with frequent transitions between swimming and non-swimming. |
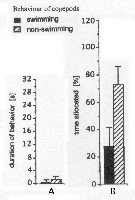 Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.211: Figs.1, 2]. As Undinula vulgaris var. tawanicus (U.v.v.t.) Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.211: Figs.1, 2]. As Undinula vulgaris var. tawanicus (U.v.v.t.)
Fig.1: A, Mean durations periods (seconds) of swimming versus non swimming behaviour by U.v.v.t. from north-east coast of Taiwan (near Pitou) in daylight in July 1993.
Fig.2: B, mean percentages of time allocated to swimming versus non-swimming behaviour by U.v.v.t. |
| | | | Loc: | | | South Africa (E & W), Angola (Baia Farta), Congo, Sao Tomé and Principe Is., Angola, Atlant. (central equatorial), Congo, G. of Guinea, off Lagos, Ivorian shelf, Dakar, Cape Verde Is., off NW Cape Verde Is., Morocco-Mauritania, W Azores, New York, Long Island, off Woods Hole, off S Nova Scotia, Argentina, Brazil (off Rio de Janeiro, Campos Basin, off Vitoria-Cabo de Sao Tomé, off Macaé, Camamu, off Natal, off Amazon), Barbados Is., Caribbean Colombia, Bahia de Mochima (Venezuela), Cariaco Basin, Caribbean Sea, Jamaica (Discovery Bay), Yucatan, E Costa Rica, Gulf of Mexico, off W Mississipi River, Straits of Florida, Florida, off Miami, Sargasso Sea, Station "S" (32°10'N, 64°30'W), off Bermuda, Gulf Stream (41°N,58°W), E Medit. (Ionian Sea-Cyprus), G. of Aqaba, Red Sea, G. of Aden, Arabian Gulf, Zanzibar Channel, Somalia, Arabian Sea, Maldive Is., Natal, Zanzibar region, Madagascar (Nosy Bé), Rodrigues Is. - Seychelles, Mascarene Basin, India (Goa, Saurasthra coast, off Cochin, Madras, Gulf of Mannar, Palk Bay, Godavari region, Porto Novo, Kakinada Bay, Lawson's Bay), Bay of Bengal, Andaman Sea (Batten Island), W Malay Peninsula (Andaman Sea), W Australia, Straits of Malacca, Indonesia-Malaysia, Sarawak: Bintulu coast, SW, Java (Cilacap Bay, Jakarta Bay-Seribu Islands, Lombok Sea, Flores Sea, Ambon Bay, Tioman Is., N Celebes, Thaïland, Philippines, N Mindanao (Butuuan Bay), Viet-Nam (Cauda Bay), G. of Tonkin, Hong Kong, China Seas (Yellow Sea, East China Sea, South China Sea, E Kuroshio Current), Taiwan Strait, Taiwan (S, E, SW, W, NW, off Danshuei River, N, Mienhua Canyon, NE, Pitou, Kueishan Is.), Okinawa, S Korea, Korea Strait, Japan Sea, Tsushima Straits, Japan (Kuchinoerabu Is., Ariake Bay, Suruga Bay, Tokyo Bay), Bering Sea, Palau Is., Pacif. W (equatorial), Australia (G. of Carpentaria, Great Barrier, Brisbane River estuary, Moreton Bay, Shark Bay), New Caledonia, Moorea Is. (lagon), Tuamotu, Is. Ahe atoll lagoon, Bikini Is., Hawaii, off S Hawaii, Kaneohe Bay, off California, BaJa California (Bahia Magdalena, W), Gulf of California, La Paz, W Mexico, Acapulco Bay, G. de Tehuantepec, W Costa Rica, W Panama, Clipperton Is., Bahia Cupica (Colombia), Galapagos-Ecuador, Chile, Tuamotu Archipelago (Ahe atoll, Tikehau atoll ) | | | | N: | 345 | | | | Lg.: | | | (14) F: 2,8-2,1; M: 2,6-2,13; (28) M: 2,5-2,08; (34) F: 3,05-2,4; M: 2,5-2,25; (45) F: 2,85-2,4; M: 2,5-2,25 ; (47) F: 2,8-2,4; M: 2,45-2,3; (73) F: 2,99-2,66; M: 2,72-2,36; (81) F: 2,3-1,9; M: 2,3-1,9; (101) M: 2,04; (104) F: 2,8; M: 2,25; (105) F: 2,83; (125) F: 2,81-2,75; M: 2,69-2,31; (137) M: 2,9; (150) F: 3-2,62; M: 2,52-2,17; (196) F: 2,95; (237) F: 2,5; M: 2; (249) F: 2,7-2,52; M: 2,41-2,18; (334) F: 2,6-1,8; M: 2,3-2; (290) F: 2,95-2,8; M: 2,65-2,4; (530) F: 2,2; M: 2,1; (785) F: 2,86-2,58; M: 3,23-2,48; (786) F: 3-2,8; M: 2,6-2,47; (795) F: 2; (991) F: 2,25-3,25; M: 2,04-2,5; (1122) F: 3,0; M: 2,5; (1130) F: 2,574; M: 2,223; (1230) F: 2,5-3,1; M: 2,1-2,6; {F: 1,80-3,25; M: 2,00-3,23} | | | | Rem.: | Epi-mésopélagique. Oceanic.
Peut-être lié aux effets de terre. Sargasso Sea: 0-500 m.
Undinula vulgaris giesbrechti Sewell,1929 (F)
Syn.: Undinula vulgaris major Vervoort, 1946 (p.73); Tanaka, 1960 (p.20); Wickstead, 1963 (p.293: parasite)
Ref.: Sewell, 1929 (p.31); Vervoort, 1946 (p.73, Rem.); Krishnaswamy, 1953 (p.109); Brodsky, 1962 c (p.105, figs.F); Kasturirangan, 1963 (p.13, fig.F); Vervoort, 1963 b (p.84); Silas, 1972 (p.644); Brodsky, 1972 (1975) (p.120); Greenwood, 1976 (p.9, fig.F); Zheng Zhong & al., 1984 (1989) (p.228, figs.F, M); Fleminger, 1986 (p.92, figs.F)
Loc.: Australie (Baie Moreton), Indonésie, Zanzibar Channel, Inde, Caraïbes, Pacif. NW
Lg.: (28) F: 3-2,5; (66) F: 2,81-2,52; M: 2,41; (795) F: 2,5; M: 2
Undinula vulgaris plumosus (Wolfenden,1906) (F)
Syn.: Calanus vulgaris plumosus Wolfenden, 1905 (1906) (p.994, figs.F)
Ref.: Sewell, 1912 (p.353, 356); 1929 (p.31); 1948 (p.407); Silas, 1972 (p.644).
Undinula vulgaris typica Sewell,1929 (F,M)
Syn.: Undinula vulgaris minor : Sewell, 1929 (p.37); Tanaka, 1960 (p.20)
Ref.: Sewell, 1929 (p.31); Vervoort, 1946 (p.73, Rem.); Brodsky, 1962 c (p.105, fig.F); Kasturirangan, 1963 (p.13, fig.F); Vervoort, 1963 b (p.84, Rem.); Björnberg, 1963 (p.16); Silas, 1972 (p.644); Brodsky, 1972 (1975) (p.120); Patel, 1975 (p.659); Greenwood, 1976 (p.9, fig.F), Goswami & al., 1977 (tab.3); Sazhina, 1985 (p.30, figs.Nauplius); Fleminger, 1986 (figs.F, p.92)
Loc.: G. de Mannar, Is. Seychelles, Inde W
Lg.: (28) F: 3,05-2,25; (29) F: 2,189-1,868; (66) F: 1,86; M: 1,87-1,84
Undinula vulgaris zeylanica Sewell,1929 (F)
Syn.: Undinula vulgaris major : Sewell, 1929 (p.37)
Ref.: Sewell, 1929 (p.32, 37); Vervoort, 1946 (p.73, Rem.); Brodsky, 1962 c (p.105, fig.F); Kasturirangan, 1963 (p.13, Rem.); Silas, 1972 (p.644); Brodsky, 1972 (1975) (p.120); Fleminger, 1986 (p.92, figs.F)
Loc.: G. de Mannar
Lg.: (28) F: 2,75-2,35.
Undinula vulgaris giesbrechti Sewell
Ref.: Patel, 1975 (p.659)
? Undinula vulgaris var. taiwanicus : Hwang & Turner, 1995 (p.207, figs.1, 2, 3, table table 1, 2, Rem: behavoriour)
Loc.: NE coast of Taiwan (25°N, 122°E).
Voir aussi les remarques en anglais | | | Dernière mise à jour : 25/10/2022 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 15 juin 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
