|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Clausocalanoidea ( Superfamille ) |
|
|
|
Euchaetidae ( Famille ) |
|
|
|
Euchaeta ( Genre ) |
|
|
| |
Euchaeta marina (Prestandrea, 1833) (F,M) | |
| | | | | | | Syn.: | Cyclops marinus Prestandrea, 1833;
Euchaeta prestandreae Philippi, 1843; Claus, 1863 (p.183, 185, figs.F,M); Thompson, 1888 d (p.140); no Brady, 1883 (p.60); no ? King & Hida, 1955 (p.11);
E. communis Dana, 1849; Dahl, 1894 c (p.12, Rem.);
Euchäta marina : Giesbrecht, 1892 (p.262, 772, figs.F,M);
E. atlantica Lubbock, 1856;
E. sutherlandii Lubbock, 1856;
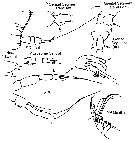
no E. marina : A. Scott, 1909 (p.67); Mori, 1937 (1964) (p.44, figs.F,M); Dakin & Colefax, 1940 (p.99, figs.F,M); Krishnaswamy, 1953 (p.117); ? King & Hida, 1955 (p.11); Yamazi, 1958 (p.148, Rem.); Grice, 1962 (p.199, fig.F); ? Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); Anraku & Azeta, 1965 (p.13, Table 2, fish predator); Tanaka & Omori, 1968 (p.220); ? Morris, 1970 (p.2300); Itoh, 1970 (p.4, Tab.1); Gueredrat, 1971 (p.300, fig.2, 10, Table 1, 2); ? Subbaraju & Krishnamurphy, 1972 (p.25, 26 ); Timonin, 1976 (p.79); Greenwood, 1977 (p.57, figs.F, Rem.); Chen Q-c, 1980 (p.795); Zheng Zhong & al., 1984 (1989) (p.237, figs.F,M); ? Williams D. & al., 1988 (p.580); ? Yoo, 1991 (tab.1); Arinardi, 1991 (p.295); He & al., 1992 (p.250); ? Shih & Young, 1995 (p.70); ? Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Takenaka & al., 2012 (p.1669, fig.2, 3, Table 1, bioluminescence assay); ? Noda & al., 1998 (p.55, Table 3, occurrence); | | | | Ref.: | | | Giesbrecht & Schmeil, 1898 (p.38, Rem. F,M); I.C. Thompson, 1903 a (p.18); Thompson & Scott, 1903 (part., p.234, 244); Wolfenden, 1911 (p.299); ? Sewell, 1912 (? part., p.353, 360); ? Pesta, 1912 a (p.45); ? Sewell, 1914 a (p.215); Pesta, 1913 (p.32); 1920 (p.512); Sars, 1925 (p.104); Farran, 1929 (p.208, 237); Rose, 1929 (p.24); ? Sewell, 1929 (p.148); Sciacchitano, 1930 (p.15, figs.F,M); Pesta, 1943 (p.17, figs.M); Lysholm & al., 1945 (p.21); Sewell, 1947 (p.113, figs.F, Rem.); Brodsky, 1950 (1967) (p.198, figs.F,M); Marques, 1953 (p.99); 1958 (p.210); Heinrich, 1961 b (p.93); Vervoort, 1957 (p.76, Rem.); Chiba & al., 1957 (p.308); 1957 a (p.11); Fish, 1962 (p.13); Gaudy, 1962 (p.93, 99, Rem.: p.106, Pl.IV, Tableau 5: development: juv.); 1963 (p.23); Paiva, 1963 (p.41); Vervoort, 1963 b (part. p.158, Rem.); M. Bernard, 1964 (p.52, figs.N); Crisafi, 1965 b (p.263, figs.F,M, juv.); ? Saraswathy, 1966 (1967) (p.77); Owre & Foyo, 1967 (p.54, figs.F,M, Rem.); Mazza, 1967 (p.145, 161, F,M, juv.); 1968 (fiche CIESMM n°1); Ramirez, 1969 (p.63, figs.F,M); Shih & al., 1971 (p.39); Razouls, 1972 (p.94, Annexe: p.48, figs.F,M); Björnberg, 1972 (p.26, Rem.N, figs.); Kos, 1972 (Vol. I, figs.F, M, Rem.); Tanaka, 1973 (part., p.134, figs.F, Rem.); Marques, 1973 (p.238); 1974 (p.15); Bradford, 1974 (p.159, figs.F,M, fig.7, Rem.); Park, 1975 c (p.5, figs.F,M); ? Marques, 1976 (p.990); ? Tranter, 1977 (p.596); Björnberg & al., 1981 (p.633, 635, figs.F,M); ? Marques, 1982 (p.758); Sazhina, 1982 (p.1158, Rem., fig. N); 1985 (p.47, figs. N); Bradford & al., 1983 (p.18); Baessa de Aguiar, 1987 (p.41, figs.F, Rem.); Blades-Eckelbarger, 1991 (p.250, figs.M); Park, 1995 (p.15, Redescr.F,M, figs.F,M); Hure & Krsinic, 1998 (p.101); Mauchline, 1999 (n°182, p.8, figs.F,M); Bradford-Grieve & al., 1999 (p.880, 925, figs.F,M); Bragina, 1999 (p.195); Braga & al., 1999 (p.84, 86, 89, Rem.: Biol. mol., Table 1 , figs.6, 7, 8); G. Harding, 2004 (p.52, figs.F,M); Mulyadi, 2004 (p.71, figs.F,M, Rem.); Conway, 2006 (p.20, copepodides 1-6, Rem.); Avancini & al., 2006 (p.85, Pl. 54, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.649, figs.F,M, Rem.); Phukham, 2008 (p.119, figs.F) | 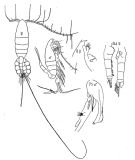 issued from : Giesbrecht in M. Rose in Faune de France. 26- Copépodes pélagiques, Paul Lechevalier, Paris, 1933; [Fig.93, p.113]. Female and Male
|
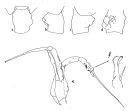 issued from : J.M. Bradford in Pacific Science, 1974, 28 (2).[Fig.4, p.165; Fig.6, p.166]. Female: genital segment: a, dorsal view; b, left side; c, right side; d, ventral view. Male: a, P5; b, serrate lamella of left P5 exopod segment 2.
|
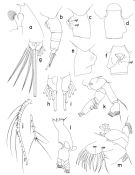 issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. California, San Diego, 1995, 29. [p.111, Fig.1]. Female: forehead (left side); b, urosome (left side); c, genital somite (left side); d, idem (dorsal); e, idem (right side); f, idem (ventral); g, caudal rami (dorsal); h, left caudal ramus (dorsal); i, idem (ventral); j, A1 (cut between segments 20 and 21); k, A2; l, Md; m, Mx1 (first inner lobe separated, posterior). agf = anterior lobe of genital flange; ase = anterior seta; pgf = posterior lobe of genital flange; pse = posterior seta. Nota: Laterally, forehead produced anteriorly into a large conical frontal eminence; Rostrum large pointing in an anteroventral direction, only slightly curved backward. Distal margin of ptosome rounded; dorsally, posterolateral corners of prosome symmetrical and somewhat angular. Genital somite in dorsal view strongly asymmetrical, with a large lobular outgrowth on right side and a small notch on left side. Laterally, right genital flange large, weakly bilobed; left genital flange smaller than right, clearly divided into a broad anterior and a narrow posterior lobe. Ventrally, posterior lobe of left genital flange extending mediad for only a short distance, leaving a relatively wide gap between it and medial edge of opposite genital flange. Within this gap and posterior to genital opening are seen conspicuous cuticular ridges. 4 distal marginal setae of caudal ramus about as long as urosome; appendicular caudal setae about 1.3 times length of body. A1 extending beyond distal end of prosome by it last 3 segments. In A2, outer margin of 2nd exopodal segment strongly bulging. Inner marginal seta of last exopodal segment well developed. In Md, endopod reaching about middle of exopod. In Mx1, 1st inner lobe with 1+9+1 (1 anterior + 9 marginal + 1posterior) setae; 1st endopodal segment with 1 seta, outer lobe with 5 setae.
|
 issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. California, San Diego, 1995, 29. [p.112, Fig.2]. Female: a, Mx2 (5th lobe and endopod separated, posterior); b, Mxp (coxa posterior, the rest anterior); c, P1 (anterior); d, P2 ( (anterior); e, P3 (anterior); f, P4 (anterior). P1 = first leg; P2 = second leg; P3 = third leg; P4 = fourth leg. Nota: In Mx2, 2 endopodal setae armed with long spinules in addition to very short spinules. Basis of Mxp armed with long spinules in addition to rows of very small spinules along proximal half of medial margin. Proximal, compound exopodal segment of P1, formed of first unseparated segments, with nearly straight outer margin and well-developed outer spine reaching about 2/3 way to base of following outer spine. In P2 exopod, 2nd outer spine short of reaching base of following outer spine; 4th outer spine largest, a litthle short of reaching base of following outer spine; marginal lobe bearing 4th outer spine separated from segment by a deep and wide incision; marginal lobes carrying 2nd and 3rd outer spines separated from respective segments by a relatively deep cleft. In P3 and P4, all outer spines of 3rd exopodal segment equally small.
|
 issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. California, San Diego, 1995, 29. [p.113, Fig.3]. Male: a, forehead (left side); b, last pedigerous and genital somites (left side); c, caudal rami (ventral); d, left caudal ramus (lateral); e, A2; f, Md; g, Mx1 (posterior); h, Mxp (coxa posterior, the rest anterior); i, first leg (anterior); j, second leg (anterior); k, 5th pair of legs (anterior); l, exopod of left 5th leg (anterior, tilted clockwise); m, idem medial, titled counterclockwise; n, distal end of serrated lamella (anterior). dgp = digitiform process; htb = hairy tubercle; tsl = poorly sclerotized lobe.
|
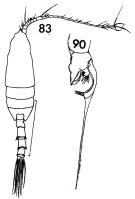 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.64, Lam. XII, figs.83, 90 ]. Male (from off Mar del Plata): 83, habitus (dorsal); 90, left P5 (detail of terminal exopodal segment) Scale bars in mm: 1.5 (83).
|
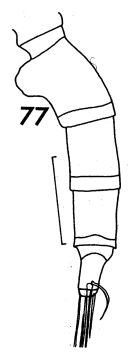 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.62, Lam. XI, fi.77]. Female (from off Mar del Plata): 77, urosome (lateral left side). Scale bar in mm: 0.4.
|
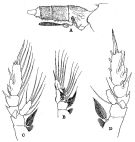 issued from : R.B.S. Sewell in The John Murray Expedition, 1933-34, Scientific Reports, VIII (1), 1947. [p.114, Fig.25]. Female (from Arabian Sea): A, urosome with spermatophore (lateral right side); B, P1; C, P2 (in small form); D, P2 (in large form). Nota: The proportional lengths of the various segments of the body (cephalon to caudal rami) in small example (3.0 mm) as 319:115:84:84:89:120:68:53:26:42 = 1000. In large example (3.683 mm) as 307:118:90:77:95:127:68:50:27:41 = 1000. There are also slight differences in the proportional lengths of the segments of A1; Differences can also be detected in the relative length of the 2nd spine on the external margin of the 3rd segment of the exopod of P2. In one specimen the joint between the 1st and 2nd segments of the exopod of P2 was incomplete, and the part of the combined segment that represented segment 1 was devoid of an inner seta. The number of ova into egg-sacs was from 7 to 15 (mean 11), in every case the ovigerous female was also bearing one or more spermatophores (the maximum number observed on any single female was 11).
|
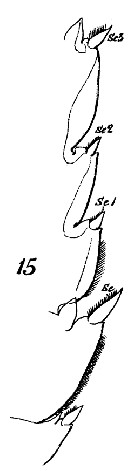
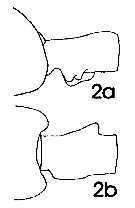 issued from : J. Mauchline in ICES Ident. Leafl. Plankton, 1999, N°182. [p.3, Fig.2: 2a-2b]. Female (Northeast Atlantic): a, genital double-somite (left side); b, same (dorsal).
|
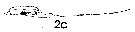 issued from : J. Mauchline in ICES Ident. Leafl. Plankton, 1999, N°182. [p.4, Fig.3: 2c]. Male (Northeast Atlantic): 2c, terminal two segments, exopodal segments of left P5.
|
 issued from : G. Harding in Key to the adullt pelagic calanoid copepods found over the continental shelf of the Canadian Atlantic coast. Bedford Inst. Oceanogr., Dartmouth, Nova Scotia, 2004. [p.52]. Female & Male. Nota: Female with deep incision, in lateral view) between frontal organ and rostrum (arrowed). Two of the six apical setae on Mx2 bearing long spines in addition to short spinules (arrowed).
|
 issued from : T. Park in Smiths. Contr. Zool., 1975, 196. [p.5, Fig.1]. Female (G. of Mexico): a, forehead (lateral); b, genital segment (ventral). Male: c, forehead (lateral); d, exopod of left P5 (anterior).
|
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1967, 1, Crustacea, 1: Copepoda. [p.55, Figs.332-335]. Female (from Florida Current): 332, Mx2. Male: 333, right P5; 334, left P5; 335, terminal segments of left P5.
|
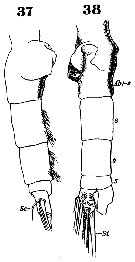 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.37, Figs.37, 38]. As Euchäta marina. Female: 37, urosome (lateral); 38, same (ventral).
|
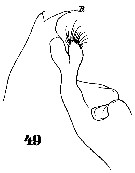 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.37, Fig.49]. As Euchäta marina. Female: 49, forehead (lateral). R = rostrum.
|
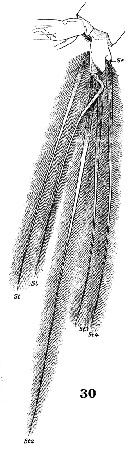 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.37, Fig.30]. As Euchäta marina. Female: 30, caudal rami and setae (ventral). Nota: Se = outer seta; St = terminal seta.
|
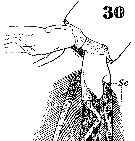 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.37, Fig.30]. As Euchäta marina. Female: 30, caudal rami (ventral; magnified).
|
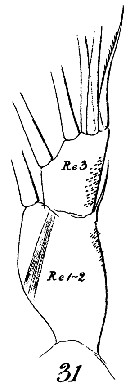
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.30]. As Euchäta marina. Female: 30, A1 (segments 1 to 18; ventral view). S di = distal seta.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.30]. As Euchäta marina. Female: 30, A1 (segments 17 to 24-25; ventral view). S di = distal seta; S pr = proximal seta.
|
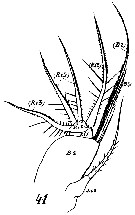
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Figs.1, 2]. As Euchäta marina. Female: 1, A2 (posterior view); 2, A2 (anterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.41]. As Euchäta marina. Female: 41, Mx1 (anterior view). B2 = basis; Li 2 = inner lobe 2.; Ri = endopod.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.28]. As Euchäta marina. Female: 28, Mx2 (posterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.8]. As Euchäta marina. Female: 8, Mxp (B1 anterior view, rest posterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.15, Fig.31]. As Euchäta marina. Female: 31, exopod of P1 (posterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.46]. As Euchäta marina. Female: 46, exopodite 2 and 3 of P2 (posterior view)
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.15]. As Euchäta marina. Female: 15, exopodal outer margin of P3 (posterior view). Se = outer seta.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.22]. As Euchäta marina. Female: 22, exopodal outer margin of P4.
|
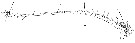 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.29]. As Euchäta marina. Male: 29, A1 (ventral view). Ae = aestherasc; S di = distal seta; Spr = proximal seta.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Figs.16, 17]. As Euchäta marina. Male: 16, right P5 (posterior view); 17, left P5.
|
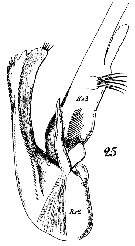 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.25]. As Euchäta marina. Male: 25, exopodite 2 and 3 of left P5.
|
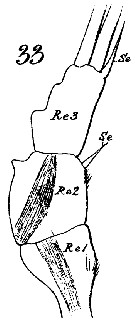 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.15, Fig.33]. As Euchäta marina. Male: 33, exopod of P1 (posterior view).
|
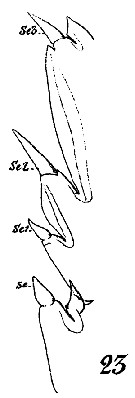 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.23]. As Euchäta marina. Male: 23, outer margin of exopodite 2 and 3 of P2.
|
 issued from : P.E. Lapernat & C. Razouls in Vie Milieu, 2002, 52 (1). [p.28, Pl. VI, fig.10]. Masticatory edge of Md gnathobase female (from off Malta, Mediterranean Sea). Nota: Itoh's index: 858.9 (number of teeth: 7).
|
 issued from : P.E. Lapernat & C. Razouls in Vie Milieu, 2002, 52 (1). [p.21, Pl. II, fig.6]. Masticatory edge of Md gnathobase female (from off Malta, Mediterranean Sea). Nota: The Itho's index vale for the the mandibular gnathobase = 858.9.
|
 issued from : R. Gaudy in Rev. Trav. St. Mar. End., Bull. 27 (42). [p.128, Tableau 5]. Identification of copepodids stages Euchaeta marina. A: Stage; B: number of swimming legs; C: number of thoracic segments; D: number of abdominal segments; E, Size.
|
 issued from : N. Phukham in Species diversity of calanoid copepods in Thai waters, Andaman Sea (Master of Science, Univ. Bangkok). 2008. [p.212, Fig.86]. Female (from W Malay Peninsula):a, habitus (dorsal); b, forehead (lateral); c-d, genital segment (lateral and dorsal, respectively); e, P2. Body length after drawing: F = 3.375 mm.
|
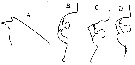 issued from : O. Tanaka in I O B C Handbook, 1973, IV. [p.134, Fig.4]. With doubt. Not taken into account by Park (1995, p.15)). Female (from Indian Ocean); A, forehead (lateral); B-D, genital segments for different specimens. Nota: There is some doubt concerning a good identification of this species because possible confusion with the allied forms E. marinella and E. rimana.
|
 issued from : O. Tanaka in I O B C Handbook, 1973, IV. [p.148, Fig.9: 1]. With doubt. Not taken into account by Park (1995, p.15). Female: 1, genital segment (dorsal). Nota: Genital segment asymmetrical, produced on the right side about the middle.
|
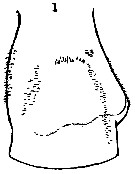
 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.71, Fig.40]. Female (from Indonesian Seas): a, habitus (dorsal); b, forehead (lateral); c, posterior part of last thoracic segment and genital complex (lateral right side); d, P2. Male: e, habitus (dorsal); f, forehead (lateral); g, posterior part of last thoracic segment and genital somite (lateral left side); h, P5; i, exopod of left P5.
|
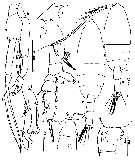
 Issued from : T. Park in Smithsonian Contr. Zool., 1975, 196. [p.5, Fig.1]. Female (from G. of Mexico & Caribbean Sea): a, forehead (lateral); b, genital segment (ventral). Male: c, forehead; d, exopod of left P5 (anterior). Nota: In the Gulf of Mexico and western Caribbean Sea, this species is the most common euchaetid in the epipelagic zone; although its batymetrical distribution extends occasionally down to a depth of about 500 m, its main concentration usually appears between depths of 25 and 100 m.
|
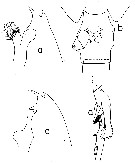
 Issued from : T. Park in Antarctic Res. Ser., 1978, 27 (4). [p.206, Fig.65]. Female (from 26°36'S, 36°41'W & 36°42'S, 53°09'W): A, forehead (lateral; B-E, distal end of metasome and genital segment (dorsal, left side, right side, and ventral, respectively); F, outer lobe of Mx1; G, P1 (anterior); H, P2 (anterior).
|
 Issued from : T. Park in Antarctic Res. Ser., 1978, 27 (4). [p.206, Fig.65]. Male (from 26°36'S, 36°41'W & 36°42'S, 53°09'W): A, forehead (lateral); B, last metasomal and genital segment (lateral); C, P5 (anterior); D, middle part of exopod of left P5 (anterior); E, middle part of exopod of left P5 (medial). Nota : Proportional lengths of prosome and urosome about 70 : 30 : Forehead similar to that of female, but frontal eminence pointing downward and rostrum curved backward. Last metasomal segment prolonged posteriorly on each side into lappet covering about ½ of genital segment; Laterally, posterodorsal margin bearing toothlike process. P5 biramous left and right. Right endopod slightly longer 1st exopodal segment. Left endopod rudimentary. Distal exopodal segment of right leg tapering into spiniform process longer than rest of leg. 2nd segment of left exopod with elongated hollow serrated lamella, slender digitiform process, and small conical tubercle. 3rd segment of left exopod a little shorter than rest of leg, consisting of hollow proximal portion and long tapering distal part ; tuft of spinules and digitiform process present at junction of these two parts.
|
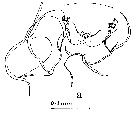 issued from : J.M. Bradford in Pacific Science, 1974, 28 (2). [p.166, Fig.5, a]. Female: c, genital field and right border of genital segment. Nota: Genital field in ventral view with genital pads not meeting in midline, gap between them about 1/2 width of right pad.
|
 Euchaeta marina Euchaeta marina Female: 1 - See Key to marina species Group. 2 - Laterally, genital somite less than 1.5 times as long as deep (Fig.1-b). 3 - In P2 exopod, 2nd outer spine of 3rd segment longer than outer spine of 2nd segment (Fig.2-d). 4 - Ventrally, genital field with wide gap between genital flanges (Fig.1-f).
|
 issued from : J.M. Bradford in Pacific Science, 1974, 28 (2). [p.164, Fig.2]. Female (from 28°35'N, 73°33W): Mx1.
|
 issued from : J.M. Bradford in Pacific Science, 1974, 28 (2). [p.164, Fig.3]. Comparison between exopods of P1 and P2 female of Euchaeta marina (a, b), Euchaeta rimana (c, d) and Euchaeta marinella (e, f). Nota: Exopod of P1: Bb ± 2/3 BC, Cc ± 4/3 BC. Exopod of P2: Aa ≤ AB, Bb ± 1/2 BC, Cc ≤ CD. In E. rimana P1 exopod: Bb ±2/3 BC, Cc ≥ BC. In E. marinella exopod of P1: Bb ≥ 1/2 BC, Cc ≥BC; exopod of P2: Aa ± 3/4 AB, Bb ≤ 1/2 BC, Cc ≤ 1/2 CD.
|
 issued from : J.M. Bradford in Pacific Science, 1974, 28 (2). [p.167, Table 2] Female and Male: comparison between Euchaeta marina and allied forms.
|
 Issued from : M.S. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. After Brodsky, 1962. Female: 1, habitus (dorsal) with eggss; 2, habitus (lateral); 3, forehead (lateral); 4, urosome (lateral); 5, genital segment; 6, P2; 7, P1. Male: 8, habitus (lateral); 9, distal end of exopod (serrated lamelle nA) of P5.
|
 Issued from : G. Belmonte in The Eur. Zool. J., 2018, 85 (1). [p.274, Fig.1]. Euchaeta marina (Prestandrea, 1833). a, first nauplius; b, metanauplius; c, Mxp; d, extremity of A1- d1 female, d2 male; c, 5th pair of thoracic legs, male; f, caudal ramui- f1 female, f2 male; g, adult male (lateral view); h
, adult female (lateral view). (Redrawn from different sources: a from Sazhina 1985; b from Bjornberg, 1972; c-g from Giesbrecht 1892; h: from Claus, 1863.
| | | | | Ref. compl.: | | | Cleve, 1904 a (? part., p.190); Pearson, 1906 (p.16, Rem.); ? Cazrl, 1907 (p.15); Rose, 1925 (p.152); Massuti Alzamora, 1942 (p.90); Wilson, 1942 a (part., p.185); Oliveira, 1945 (p.191); Wilson, 1950 (part. p.214); Deevey, 1960 (p.5, Table II, annual abundance); ? Fagetti, 1962 (p.21); ? Ganapati & Shanthakumari, 1962 (p.8, 15); Cervigon, 1962 (p.181, tables: abundance distribution); Grice & Hart, 1962 (p.287, table 3); V.N. Greze, 1963 a (tabl.2); Gaudy, 1963 (p.23, Rem.); Björnberg, 1963 (p.37, Rem.); Unterüberbacher, 1964 (p.23); ? De Decker & Mombeck, 1964 (p.12); Shmeleva, 1965 b (p.1350, lengths-volume-weight relation); ? Mullin, 1966 (p.546, Table I, III, diet); Dawson, 1966 (p.176); Pavlova, 1966 (p.43); Neto & Paiva, 1966 (p.23, Table III); Chakroun, 1966 (p.67, Table); ? Furuhashi, 1966 a (p.295, vertical distribution in Kuroshio region, Table 9); Mazza, 1966 (p.70); 1967 (p.367); Ehrhardt, 1967 (p.738, figs.12, 51-53, geographic distribution, Rem.); ? Grice & Hulsemann, 1967 (p.16); ? Fleminger, 1967 a (tabl.1); ? Itoh, 1970 (tab.1); Vinogradov, 1968 (1970) (p.78, 80, 278); Evans, 1968 (p.13); Dowidar & El-Maghraby, 1970 (p.268); Park, 1970 (p.476); Deevey, 1971 (p.224); Carli, 1971 (p.373, tab.2); Salah, 1971 (p.319); ? Timonin, 1971 (p.281, trophic group); Roe, 1972 (p.277, tabl.1, tabl.2); 1972 b (p.526); Brodsky, 1972 (P.256); Apostolopoulou, 1972 (p.327, 347); Binet & al., 1972 (p.71); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night, Table II:%, Table IV: thermal stability vs upwelling); Ibanez & Seguin, 1972 (p.81, annual cycle, multivarite analysis); ? Heinrich, 1973 (p.95); ? Björnberg, 1973 (p.329, 386); ? Patel, 1975 (p.659); Gaudy, 1975 (p.109, fig.1, Table I, 4, respiration); Ikeda, 1976 (p.51, respiration rate); ? Frontier, 1977 a (p.14); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); ? Timonin & Voronina, 1977 (p.290, fig.7); Carter, 1977 (1978) (p.35); Arashkevich, 1978 (p.118, Table: diets); ? Stephen & Iyer, 1979 (p.228, tab.1, 3, 4, figs.3, 4); Dessier, 1979 (p.201, 205); Vaissière & Séguin, 1980 (p.23, tab.2); Svetlichnyi, 1980 (p.28, Table 1 as Fuchaeta marina, passive submersion); Sreekumaran Nair & al., 1981 (p.493, fig.2cont.); Madhupratap & al., 1981 (p.262, Table 2: abundance vs. thermocline); Madhupratap & al., 1981 (p.266, fig.2j: abundance vs. geographic transect); Kovalev & Shmeleva, 1982 (p.83); Kovalev & Shmeleva, 1982 (p.83); Gaudy & Boucher, 1983 (p.37, Table 1, 2, 3, 5, figs.1, 2, Rem.: metabolism); Minkina, 1983 (p.38, speed swimming); ? Guangshan & Honglin, 1984 (p.118, tab.);Tremblay & Anderson, 1984 (p.5); De Decker, 1984 (p.316, 349: chart); Cummings, 1984 (p.163, Table 2); Scotto di Carlo & al., 1984 (p.1042); Kinzer & Schulz, 1985 (tab.4); Regner, 1985 (p.11, Rem.: p.31); Longhurst, 1985 (tab.2); Brenning, 1985 a (p.23, Table 2); Sazhina, 1985 a (p.491, tab.3); Petipa & Borichenko, 1985 (tab.1,2); Moraitou-Apostolopoulou, 1985 (p.303, occurrence/abundance in E Mediterranean Sea); Jansa, 1985 (p.108, Tabl.III, V); Sarkar & al., 1986 (p.178); Shuert & Hopkins, 1987 (p.49); Shuert & Hopkins, 1987 (p.30, feeding: diet composition); Renon, 1987 (tab.2); Gorsky & al., 1988 (p.133, Table I, C-N composition); Lozano Soldevilla & al., 1988 (p.58); Hernandez-Trujillo, 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Herman, 1989 (p.247); Mitra & al., 1990 (fig.3); Suarez & al., 1990 (tab.2); Othman & al., 1990 (p.565, Rem.); Gajbhiye & al., 1991 (p.189); Lindo, 1991 (tab.3); Suarez & Gasca, 1991 (tab.2); Hernandez-Trujillo, 1991 (1993) (tab.I); Samba Diouf, 1991 (p.104); Huntley & Lopez, 1992 (p.201, Table B1, growth rate, temperature-dependent production); Suarez, 1992 (App.1); Seguin & al., 1993 (p.23); Hays & al., 1994 (tab.1); Kouwenberg, 1994 (tab.1); Roff & al., 1995 (p.165, Table 1, 3, 5, nauplii development times, nauplii copepodites growth rates, naupllii vs. bacteriophagy/picoplankton, naupliar production); Webber & Roff, 1995 a (p.481, table 1, 2, 3, 6, 7, 8, biomass, production); Palomares Garcia & Vera, 1995 (tab.1); ? Kotani & al., 1996 (tab.2); Webber & al., 1996 (tab.1); Suarez-Morales & Gasca, 1997 (p.1525); Sharaf & Al-Ghais, 1997 (tab.1); ? Go & al., 1997 (tab.1); ? Hwang & al., 1998 (tab.II); Gilabert & Moreno, 1998 (tab.1,2); Mauchline, 1998 (tab.63, 30, 42, 47, 64, 65); Alvarez-Cadena & al., 1998 (tab.1,2,3,4); Suarez-Morales, 1998 (p.345, Table 1); Suarez-Morales & Gasca, 1998 a (p.109); Hopcroft & al., 1998 (tab.2); Lapernat, 1999 (p.17, 55); Siokou-Frangou, 1999 (p.476); Hernandez-Trujillo, 1999 (p.284, tab.1); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); El-Serehy, 1999 (p.172, Table 1, occurrence); Lopes & al., 1999 (p.215, tab.1); Neumann-Leitao & al., 1999 (p.153, tab.2); Lapernat, 2000 (tabl.3, 4); Suarez-Morales & al., 2000 (p.751); Fernandez-Alamo & al., 2000 (p.1139, Appendix); Plounevez & Champalbert, 2000 (p.175, Table III, IV, V, abundance vs fish); ? Escribano & Hidalgo, 2000 (p.283, tab.2); Suarez-Morales & Gasca, 2000 (1247, tab.1); Suarez-Morales & al., 2000 (p.751, tab.1); Seridji & Hafferssas, 2000 (tab.1); Lopez-Salgado & al., 2000 (tab.1); Hidalgo & Escribano, 2001 (p.159, tab.2); Hidalgo & Escribano, 2001 (p.158, fig.5); Lapernat & Razouls, 2001 (tab.1); Hernandez-Trujillo & Suarez-Morales, 2002 (p.748, tab.1); Beaugrand & al., 2002 (p.179, figs.5, 6); Pusch & al., 2004 (251, tab.3); CPR, 2004 (p.54, fig.155); Kazmi, 2004 (p.229); Dias & Bonecker, 2005 (p.100 + poster); Alvarez-Silva & al., 2005 (p.39); Ashjian & al., 2005 (p.1380: tab.2); Berasategui & al., 2005 (p.313, fig.2); ; Manning & Bucklin, 2005 (p.233, Table 1); Dias Araujo, 2006 (p.49, Rem., chart); Durbin & Casas, 2006 (p.2537, Table 2b); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño, Rem.: p.70, 72); Khelifi-Touhami & al., 2007 (p.327, Table 1); Jitlang & al., 2008 (p.65, Table 1); Fernandes, 2008 (p.465, Tabl.2); McKinnon & al., 2008 (p.844: Tab.I, p.848: Tab. IV); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.513); Muelbert & al., 2008 (p.1662, Table 1); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); ? Pagano, 2009 (p.116); Miyashita & al., 2009 (p.815, Tabl.II); C.E. Morales & al., 2010 (p.158, Table 1); Hafferssas & Seridji, 2010 (p.353, Table 3); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Hidalgo & al., 2010 (p.2089, Table 2); Sun & al., 2010 (p.1006, Table 2); Dias & al., 2010 (p.230, Table 1); Mazzocchi & Di Capua, 2010 (p.425); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Fazeli & al., 2010 (p.153, Table 1); ? Maiphae & Sa-ardrit, 2011 (p.641, Table 2); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Shiganova & al., 2012 (p.61, Table 4); Uysal & Shmeleva, 2012 (p.909, Table I); Mulyadi & Rumengan, 2012 (p.202, Rem.: p.204); Miloslavic & al., 2012 (p.165, Table 2, transect distribution); Naz & al., 2012 (p.61, Table 4, relative abundance); Hidalgo & al., 2012 (p.134, Table 2) ; Belmonte & al., 2013 (p.222, Table 2, abundance vs stations); Teuber & al., 2013 (p.28, Table 1, respiration rates); Teuber & al., 2013 (p.1, Table 1, 2, 3, fig.5, abundance vs oxygen minimum zone, respiration rates, enzyme activity); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers. [see in Remarks]); Melo Jùnior & al., 2013 (p.363, Table 2, egg production); Lidvanov & al., 2013 (p.290, Table 2, % composition); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Lopez-Ibarra & al., 2014 (p.453, Table 2, biogeographical affinity); Mazzocchi & al., 2014 (p.64, Table 5, abundance); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production); Bode & al., 2015 (p.268, Table 1, chemical components, trophic level, geographic zone); Zakaria & al., 2016 (p.1, Table 1); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence); El Arraj & al., 2017 (p.272, table 2, spatial distribution); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Belmonte, 2018 (p.273, Table I: Italian zones); Abo-Taleb & Gharib, 2018 (p.139, Table 5, occurrence %); Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence); Bode & al., 2018 (p.840, Table 1, fig.5 as spp., respiration & ingestion rates); Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions, Table 5: indicator ecoregions). | | | | NZ: | 17 + 3 douteuses | | |
|
Carte de distribution de Euchaeta marina par zones géographiques
|
| | | | | | | | | | | | | | |  issued from : J.M. Bradford in Pacific Science, 1974, 28 (2). [Fig.7, p.167). issued from : J.M. Bradford in Pacific Science, 1974, 28 (2). [Fig.7, p.167).
Distribution of Euchaeta marina, E. rimanaand E. marinella based on specimens seen by the author. |
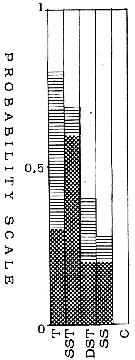 issued from : T.S.K. Björnberg in Bol. Inst. Oceanogr., Sao Paulo, 1963, XIII (1). [p.38, Fig.20]. issued from : T.S.K. Björnberg in Bol. Inst. Oceanogr., Sao Paulo, 1963, XIII (1). [p.38, Fig.20].
Probability of occurrence of Euchaeta marina in different environments.
T: Tropical waters (above 36.00 p.1000 salinity and above 20°C temperature); SST: Surface subtropical waters (salinity around 36 p.1000 and temperature 18°C or less); DST: Deeper shelf waters (salinity between 34 and 36, temperature under 20°C p.1000); SS: Surface shelf waters (same salinity and temperature above 20°C);
C: coastal waters with low salinity and variable temperature.
In each column no shading means no probability of finding the species in the samples from this environment; horizontal shading indicates the probability of finding the sepecies in percentages less than one in samples from this environment; cross shading indicates the probability of finding it in percentages higher than one and black shading represents the probability of finding it in the largest percentages of the total number of copepods.
Nota: The histogram shows this species as a copepod which prefers warm, very saline waters, therefore thermophile. As it did not occur in coastal waters it is preferentially a shelf oceanic species in Brazilian waters. It occurred in small quantities or not at all in the samples from deeper layers: therefore a surface copepod. |
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1967, 1, Crustacea, 1: Copepoda. [p.54, Table 20]. issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1967, 1, Crustacea, 1: Copepoda. [p.54, Table 20].
Vertical distribution of Euchaeta marina.
Samples collected at the 40-mile Station in the Florida Current (18 may 1958).
A: 10.37-13.27 h; B: 22.59-01.18 h. |
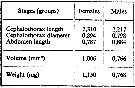 issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 20 ]. Euchaeta marina (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 20 ]. Euchaeta marina (from South Adriatic).
Dimensions, volume and Weight wet. Means for 50-60 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
 issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 33. Jahrgang 1981. Mat.-nat. wiss. Reihe, 6. [p.2, Fig.1]. issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 33. Jahrgang 1981. Mat.-nat. wiss. Reihe, 6. [p.2, Fig.1].
T-S Diagram for Euchaeta marina, E. acuta, E. hebes (= Paraeuchaeta hebes), E. paraconcinna from Cap Blanc (Mauritania).
SO: Southern Surface Water (S °/oo: 34,50; T°C: 29,0); ND: Northern Water of the Surface Layer (S °/oo: 37,5; T°C: 21,0); SD: Southern Deep Water of the surface layer (S °/oo: 35,33; T°C: 13,4). See commentary in Temora stylifera and Brenning (1985 a, p.6). |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1].
Length-weight regression of Euchaeta marina in oceanic waters of Discovery Bay (Jamaica) in 60- and 200 m vertical hauls, using 200- and 64-µm mesh aperture of plankton nets, from June 1989 to July 1991 (monthly samples). |
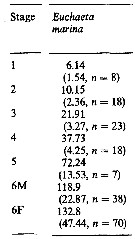 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 2]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 2].
Mean biomass (µg) of copepodite stages for Euchaeta marina from Discovery Bay (Jamaica), with standard deviation and n number of individuals measured. |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 3]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 3].
Stage weight increments (In mmax mmin-1) for Euchaeta marina from Discovery Bay (Jamaica) calculated from mean biomass of each stage. |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 4]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 4].
Stage duration (days) for copepodite stages of Euchaeta marina from Discovery Bay (Jamaica), n: number of observations. |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 5]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 5].
Mean number of eggs per female for Euchaeta marina, and corrected egg development time in days. n: number of observations. |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 6]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 6].
Estimates of generation times (in days) from egg to first egg clutch for Euchaeta marina from Discovery Bay (Jamaica), based on minimum and maximum development time of individual life history stages. Centering indicates Max and Min values identical. |
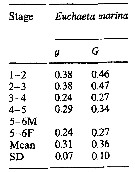 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 7]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 7].
Growth coefficient (g) and daily specific growth rate (G) for Euchaeta marina from Discovery Bay (Jamaica). |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 8]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 8].
Euchaeta marina: Minimum female weight (MFW), mass of eggs (M of E) and mass of female and eggs (MF + E) all in µg AFDW. Growth coefficient (g) and daily specific growth rate (G) for eggs.
AFDW = ash-gree dry weight. |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.483]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.483].
Equations used for the estimation of Euchaeta marina production as for other copepods in the Discovery Bay (Jamaica). |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.483]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.483].
Equations used for estimation of egg production in Euchaeta marina as for other copepods from Discovery Bay (Jamaica) |
 Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.170, Table 5]. Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.170, Table 5].
Summary observations on naupliar bacterivory on FLB (fluorescently labelled bacteria) a partir Escherichia coli (±0.7 µm3 cell volume) in Discovery Bay (Jamaica).
Fluorescence levels: 0 = no materal for observation; - = no fluorescence observed; + = low fluorescence due to few FLB observe; * = highly fluorescence due to many FLB.
The authors show of the 11 species examined (Centropages velificatus, Acartia lilljeborgi, Paracalanus aculeatus, Temora stylifera, Undinula vulgarius, Euchaeta marina, Clausocalanus spp., Onaea sppp., Corycaeus spp.), 7 showed clear evidence of feeding on FLB (see table 5), with 92% of their nauplii ingesting FLB, clearly visible in the guts of many nauplii, and usually comprisised a discrete feeding (foregut) or 'fecal' (hindgut) bolus of cells. After only 30 min, fecal pellets were being produced already filled with FLB. Presence of FLB in guts cannot be interpreted as incidental feeding or 'drinking', because 4 species failed to ingest them (Centropages velificatus, Euchaeta marina, Clausocalanus sp., Corycaeus sp.). The very earliest stages may not ingest bacteria or other particles because they may have yolk reserves (for example Temora turbinata N1 nauplii did not ingest FLB, but later stage nauplii did). |
 Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, Table 1). Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, Table 1).
Cf. explanations of these measures in Calanoides natalis from the same authors.
Compare with Paraeuchaeta gracilis, P. aequatorialis. |
| | | | Loc: | | | South Africa (W, S, E), Atlant. (tropical, sub-tropical), Namibia, Atlant. (central equatorial), Angola, Baia Farta, Congo, G. of Guinea, off Lagos, Ivorian shelf, Dakar, off NW Cape Verde Is., Morocco- Mauritania, Great Meteor Seamount, Canary Is., Azores, Gibraltar, Argentina, Uruguay (continental shelf), Brazil (S, off Macaé, off Rio de Janeiro, Campos Basin, Cabo Frio, off Vitoria-Cabo de Sao Tomé, off Natal), off Amazon, Barbada Is., Caribbean Sea, Caribbean Colombia, Bahia de Mochima (Venezuela), Cariaco Basin, Yucatan, Jamaica (Discovery Bay), Yucatan (Ascension Bay), E Costa Rica, G. of Mexico, Louisiana (off Grand Isle), Florida, Sargasso Sea, off Bermuda: Station "S" (32°10'N, 64°30'W), Delaware Bay (outside), G. of Maine, off Woods Hole, Georges Bank, G. of St. Lawrence, off S Iceland, W Ireland, off SW Ireland, Faroe Is., North Sea, Medit. (Alboran Sea, Habibas Is., Sidi Fredj coast, Algiers, Gulf of Annaba, Castellon, Baleares, Banyuls, G. of Lions, Marseille, Villefranche-s-Mer, Ligurian Sea, Tyrrhenian Sea, Messina, Gulf of Taranto, Tripoli, Malta, Adriatic Sea, Ionian Sea, Aegean Sea, Black Sea, Lebanon Basin, W Egyptian coast, Alexandria), Hurghada, N Red Sea, Gulf of Oman, ? Arabian Sea, ? Arabian Gulf, UAE coast, Karashi coast, ? Natal, ? Madagascar (Nosy Bé), ? Kavaratti & Agatti Is., ? off Cochin, ? equatorial Indian Ocean, ? India (Saurashtra coast, Porto Novo, Bombay, Lawson's bay, Hooghly estuary), E India, ? Bay of Bengal, Andaman Sea, W Malay Peninsula (Andaman Sea), Bali Sea, ? Ambon Bay, Flores Sea, Australia (North West Cape, ? G. of Carpentaria in Markina, 1972b, ? Great Barrier, Gulf of Tonkin in Brodsky, 1972 (p.256); ? Pacif. (E & W equatorial) (in Guangshan & Honglin, 1984), [ ? Korea (in Park & Choi, 1997), Yellow Sea in Sun & al., 2010; ? E China Sea (Kuchinoerabu Is.), ? Japan, ? Okinawa ], California, W Baja California (in Ahlstrom & Thrailkill, 1963); , Bahia Magdalena, W (in Lopez-Ibarra & Palomares-Garcia, 2006; Hernandez-Trujillo & al., 2010), G. of California (in Jimenez-Perez & Lara-Lara, 1988), W Mexico (in Suarez-Morales & al., 2000), G. of Tehuantepec, off W Guatemala, Costa Rica (Dome), Clipperton Is., Bahia Cupica (Colombia) ? Peru, Chile (N-S, Santiago, Concepcion) | | | | N: | 513 ? (any confusion with E. rimana in the Indian Ocean and probably in the East tropical Pacific) | | | | Lg.: | | | (3) F: 3,64-3,4; M: 3,2-2,88; (14) F: 3,3-2,73; M: 3,1-2,85; (19) F: 2,84-3,44; M: 2,88-3,4; (47) F: 3,9-2,25; M: 3,2-3,05; (73) F: 3,53; M: 3,59; (199) F: 3,57-3,19; M: 3,19-3,04; (207) F: 3,56-3,48; M: 3,52-3,3; (237) F: 3,3; M: 2,8-3,0; (244) F: 3,2-2,8; (340) M: 3,5; (432) F: 3,9-3,5; (449) F: 3,9-2,25; M: 3,2-3 ; ?(530) F: 3,8; M: 3,5; (785) F: 3,36-2,59; M: 3,18-2,56; (864) F: 2,86-3,38; (920) F: 3,33; (1111) F: 2,93-3,65; M: 2,6-3,81; (1122) F: 3,2; M: 2,45; (1230) F: 3,0-3,7; M: 3,1-3,7; {F: 2,25-3,90; M: 2,56-3,81}
The mean female size is 3.268 mm (n = 29; SD = 0.4513), and the mean male size is 3.117 mm (n = 24; SD = 0.3393). The size ratio (male : female) is 0.967 (n = 9; SD = 0.478), or ± 97 %. In samples, the sex ratio (female : male) is 1.214, but likely 1.
Prosome length: (19) F: 2,08-2,48; M: 2,12-2,36. | | | | Rem.: | épi-mésopélagique, 3000 m (off Malte).
Bradford (1974) met en doute le cosmopolitisme de cette espèce en établissant l'existence en Indonésie et dans le Pacifique de deux espèces nouvelles : E. rimana et E. marinella.
La répartition géographique de E. marina apparaît essentiellement atlantique et méditerranéenne, et sans doute dans l'océan Indien, bien que Madhupratap & Haridas (1986) ne la citent pas, contrairement à une publication de 1981 où l'espèce apparaît abondante dans la Mer des Andaman (il est à noter que les nouvelles espèces décrites par Bradford en 1974, E. marinella et rimana ne sont pas signalées). Peut-être dans le Pacifique Est bien qu'elle ne soit pas recensée par Hernandez-Trujillo & Esqueda-Escarcega, 2002 (in Appendix: p.62). Citée comme abondante par Hernandez-Trujillo & Suarez-Morales (2002) au long de la côte ouest de la Basse Californie; Les localisations dans le Pacifique nord-ouest et sud-est nécessitent aussi une confirmation sérieuse.
La présence de cette espèce est confirmée dans les mers de Chine par W. Zhang (comm. pers. 2006), comme sur les côtes du Chili (Hidalgo & al. (2012. par contre cette espèce n'est pas citée par Palorares-Garcia & al. (2018, Table 1) qui établissent la liste des copépodes dans la baie de La Paz (sud de la Basse Californie) de 1987 à 2011 qui comprend 96 espèces soit environ 21% des copépodes recenssées dans le Pacifique oriental tropical (zones 20-25 in Razouls & al. 2019). La présence de cette espèce est mentionnée par Lopez Ibarra (2008, Tables 1 and 2) mais non E. rimana un doute subsiste alors sur cette identification.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 28/10/2022 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 18 juin 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
