Introduction | Morphologie générale | Morphologie des diverses parties du corps et des appendices | Dimensions | Formes | Principales modifications structurelles | Les antennules | Les antennes |
Le cadre buccal : Labrum et labium | Les Mandibules | Les Maxillules ou premières maxilles | Les Maxilles (Maxilla) ou secondes maxilles | Les Maxillipèdes | Les pattes natatoires | Variation du nombre des segments corporels | Nomenclature des parties du corps et Métamérie | Structures et organes tégumentaires | Organes visuels |
Variations de la morphologie | Sex-ratio | Gynandromorphisme et intersexualité | Accouplement | Parthénogénèse | Caractéristique de la ponte |
Les formes larvaires | La durée du développement | La croissance | Régénération |
Anomalies | Classification et Phylogénie | Références | Photothèque
 Variation du nombre des segments corporels : Variation du nombre des segments corporels :
La segmentation apparente du corps chez les Copépodes libres paraît liée au comportement locomoteur et à l'évolution à l'intérieur des divers groupes (Kabata, 1979; Boxshall, 1985).
D'une manière générale, un comportement locomoteur plus ou moins continu, qu'il soit de nage ou de reptation, conduit à maintenir des segments au corps antérieur et postérieur plus nombreux et mieux équilibrés que chez les formes adaptées à des substrats particuliers et à nage plus intermittente.
Parmi les Calanoida, on trouve des formes chez lesquelles la céphalisation n'a pas progressé. Claus (1863) illustre ce processus dans un tableau où il montre chez les Pontellidae la région antennaire séparée.
Certaines espèces sont passées au stade suivant, chez lequel le céphalon est bien fusionné, mais où le premier segment thoracique (qui porte les maxillipèdes) a conservé son identité, comme c'est le cas chez quelques Poecilostomatoida.
Beaucoup plus commun est l'état de céphalothorax, tagme comprenant les deux segments thoraciques antérieurs fusionnés (portant les Mxp et P1).
Le Cyclopïde type illustre cette morphologie, mais on peut aussi l'observer chez les Calanoïdes et chez les Harpacticoïdes où la fusion est très générale. Le céphalothorax est alors suivi par quatre segments thoraciques individualisés portant chacun une paire de pattes. Ces segments sont de proportions inégales, mais, en règle générale diminuent en taille du second au cinquième, ce dernier étant souvent réduit (principalement chez les Cyclopoïdes) et dans une situation intermédiaire chez les Harpacticoïdes et les Monstrilloïdes.
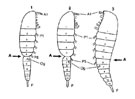 Chez de nombreux Calanoïdes, le cinquième segment thoracique fusionne avec le précédent et porte ou non une P5. Celle-ci étant toujours réduite au moins chez la femelle. Chez de nombreux Calanoïdes, le cinquième segment thoracique fusionne avec le précédent et porte ou non une P5. Celle-ci étant toujours réduite au moins chez la femelle.
Le corps présente, plus ou moins vers sa partie postérieure, une articulation marquée, considérée par les auteurs anciens (Giesbrecht, 1892; Sars, 1903; Wilson , 1932, et ultérieurement) comme un événement évolutif majeur. Cette articulation divise le corps en une partie antérieure (Prosome) et une partie postérieure (Urosome) (fig. G6). G6).
Cet emplacement, variable, mais le plus fréquemment entre les segments thoraciques 4 et 5, a conduit à l'intégration des segments ainsi séparés aux segments antérieurs et postérieurs.
Ainsi chez la plupart des Cyclopoïdes, les Harpacticoïdes et les Monstrilloïdes, le cinquième segment thoracique appartient au corps postérieur, ressemblant plus ou moins aux segments abdominaux.
Giesbrecht (1892) tire de cette situation l'existence de deux groupements: les Gymnoplea (étymol.: à abdomen nu) chez lesquels l'articulation se situe après le segment thoracique 5 (Th5), et les Podoplea (étymol.: à abdomen muni de pattes) où elle se situe après le segment thoracique 4 (Th4).
Le cinquième segment thoracique est suivi par le complexe génital; le terme de segment génital (souvent usité) doit être rejeté car il correspond à un petit tagme comprenant plus d'un segment ou somite (généralement deux). La plupart des auteurs s'accordent sur son origine somitique thoracique.
Le fait que chez les mâles de nombreux Copépodes, on observe un segment de plus que chez la femelle suggère que chez ces dernières il y a fusion de deux somites. La présence d'une paire de pattes vestigiales sur le complexe génital plaide aussi en faveur de sa nature thoracique, au moins partiellement dans sa partie antérieure. Chez les Harpacticoïdes, les deux somites du tagme génital femelle sont fusionnés, mais chez certaine on peut voir une suture dorsale. Chez le mâle, ils sont séparés.
L'abdomen proprement dit est généralement cylindrique, et présente selon les groupes un nombre de segments variable. Chez la plupart des Copépodes libres on observe trois segments chez la femelle et quatre chez le mâle, mais il n'est pas rare de constater une diminution de ce nombre par suite de l'extrême petitesse ou de la fusion du segment distal (segment anal). Chez Farranula (Poecilostomatoida) l'urosome ne montre plus qu'un seul segment chez la femelle comme chez le mâle.
Les segments abdominaux peuvent s'emboîter plus ou moins les uns dans les autres: les postérieurs à l'intérieur des précédents en raison de leur section plus réduite et l'existence d'un anneau membraneux souple antérieur à chaque segment, tandis qu'à leur extrémité postérieure on peut voir un anneau membraneux libre frangé, avec à son insertion une rangée de denticules. Cette situation de "télescopage" des segments abdominaux existe dans tous les groupes, mais est plus fréquemment rencontrée chez les Calanoïdes et les Harpacticoïdes. Ceci rend souvent difficile les mensurations de l'abdomen, surtout sur des organismes fixés et conservés, comme le comptage du nombre des segments (le segment anal peut être dans certains cas peu visible).
Le dernier segment abdominal (ou segment anal) montre un anus qui s'ouvre dorsalement, exceptionnellement à l'extrémité, recouvert par un pseudo-opercule diversement ornementé qui correspondrait au telson. Sur ce dernier segment abdominal s'articulent les rames caudales, deux sclérites plats ou subcylindriques de longueurs variées selon les espèces et pourvues de soies. Pour Bowman (1971) le terme d'uropode doit être retenu plutôt que celui de furca. L'existence d'articulations sur les soies furcales comme sur les soies des pattes natatoires confortent l'homologie (Vaupel-Klein, 1986); elles résulteraient des forces qui s'exercent au cours de la nage. Gurney (1931) cite quelques cas de structures anormales montrant une soudure des deux rames qui ressemblent alors au telson des Euphausiacés. Le débat sur la nature somitique de l'extrémité caudale (Marcotte, 1982; Mac Laughlin, 1982) montre qu'il n'y a pas unanimité sur la nature exacte de la furca.
|



 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :