Introduction | Morphologie générale | Morphologie des diverses parties du corps et des appendices | Dimensions | Formes | Principales modifications structurelles | Les antennules | Les antennes |
Le cadre buccal : Labrum et labium | Les Mandibules | Les Maxillules ou premières maxilles | Les Maxilles (Maxilla) ou secondes maxilles | Les Maxillipèdes | Les pattes natatoires | Variation du nombre des segments corporels | Nomenclature des parties du corps et Métamérie | Structures et organes tégumentaires | Organes visuels |
Variations de la morphologie | Sex-ratio | Gynandromorphisme et intersexualité | Accouplement | Parthénogénèse | Caractéristique de la ponte |
Les formes larvaires | La durée du développement | La croissance | Régénération |
Anomalies | Classification et Phylogénie | Références | Photothèque
 Les antennules (A1) : Les antennules (A1) :
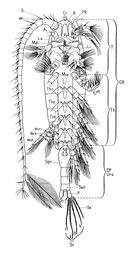

Elles sont formées d'une simple rangée de segments, plus ou moins cylindriques, décroissant en taille vers l'extrémité distale (fig. G3 , A). Le nombre de segments est variable selon les grandes coupures taxinomiques. Elles peuvent être presque identiques dans les deux sexes ou présenter chez le mâle un dimorphisme sexuel plus ou moins accentué, asymétrique ou symétrique, avec ou non une ou deux articulations vers la partie distale (fig. G3 , A, b). Des soies et aesthètes occupent des sites particuliers sur les segments et un aspect séquentiel déterminé qui permet de définir une chétotaxie (Fleminger, 1985). La fusion ou réduction des segments est analysée à partir de celle-ci (Cf in Huys et Boxshall, 1991, p.20, 362-364). Fondamentalement le nombre de segments est de 28, le segment basal porte 3 soies plus 1 aesthète, les segments 2 à 21 et 24 à 27: 2 soies + 1 aesthète (trithek), les segments 22 et 23: 1 soie + 1 aesthète, le segment distal 28: 4 soies + 1 aesthète. Dans la pratique (faunes, description) la numérotation des segments ne tient compte que des segments individualisables ou présentant une trace de séparation souvent incomplète) G3 , A). Le nombre de segments est variable selon les grandes coupures taxinomiques. Elles peuvent être presque identiques dans les deux sexes ou présenter chez le mâle un dimorphisme sexuel plus ou moins accentué, asymétrique ou symétrique, avec ou non une ou deux articulations vers la partie distale (fig. G3 , A, b). Des soies et aesthètes occupent des sites particuliers sur les segments et un aspect séquentiel déterminé qui permet de définir une chétotaxie (Fleminger, 1985). La fusion ou réduction des segments est analysée à partir de celle-ci (Cf in Huys et Boxshall, 1991, p.20, 362-364). Fondamentalement le nombre de segments est de 28, le segment basal porte 3 soies plus 1 aesthète, les segments 2 à 21 et 24 à 27: 2 soies + 1 aesthète (trithek), les segments 22 et 23: 1 soie + 1 aesthète, le segment distal 28: 4 soies + 1 aesthète. Dans la pratique (faunes, description) la numérotation des segments ne tient compte que des segments individualisables ou présentant une trace de séparation souvent incomplète)
1 - Dans le groupe des Copépodes planctoniques (vivant en pleine eau), l'antennule a un rôle essentiel(non le seul) dans la locomotion et la sustentation du fait de son allongement et de la présence de nombreuses soies plumeuses.
Chez les Calanoida, on dénombre au plus 27 segments (Erebonectes nesioticus), 25 chez Calanus finmarchicus femelle et 16 chez les femelles de Pontellopsis.
La longueur, indépendamment du nombre de segments, peut atteindre ou dépasser l'extrémité distale de la furca comme chez Calanus finmarchicus, largement les soies furcales comme chez Haloptilus longicornis femelle, ou être plus courte que le prosome comme chez Phaenna ou Euchaeta-Paraeuchaeta, voire plus courte que la tête comme chez Pseudocyclopia. Les espèces mésopélagiques semblent posséder les antennules les plus longues, alors que les espèces vivant proche des fonds les ont plus raccourcies.
Les longueurs relatives des segments dans la même espèce sont susceptibles de varier (Sewell, 1929, 1932). Pour Gaudy (1971a) la longueur de l'antennule chez Centropages typicus serait corrélée avec la densité des eaux.
La locomotion est assurée par des battements puissants d'avant en arrière, généralement discontinus. La musculature plus forte chez le mâle que chez la femelle, se traduit par des segments proximaux légèrement dilatés comme chez Calanus finmarchicus.
Les antennules du mâle peuvent être symétriques et semblables à celles de la femelle comme dans la tribu des Isokerandria comme chez Diaixis ou Stephos par exemple. Symétriques et sexuellement légèrement dimorphique, mais sans présenter d'articulation, par exemple Calanus finmarchicus comme dans la tribu des Amphascandria. Dissymétriques et sexuellement plus ou moins fortement modifié (le plus souvent à droite, mais parfois à gauche) en organe préhensile intervenant lors de l'accouplement comme dans la tribu des Heterarthrandria. Ces sections établies par Giesbrecht (1892), complétées par Sars (1901-1903), sont maintenent abandonnées comme sans valeur taxinomique (Huys et Boxshall, 1991, p.50-51).
Les antennules portent des organes récepteurs appartenant morphologiquement et fonctionnellement à deux catégories: les soies et les aesthètes (ou aesthetascs).
Les soies sont de plusieurs types morphologiques, pour l'essentiel élancées et courtes, flexibles et plus rigides, portant souvent une simple ou double rangée de fins poils leur conférant une apparence plumeuse. Elles sont supposées être des récepteurs mécaniques, certaines intervenant dans la portance notamment pour les plus longues et plumeuses, situées souvent sur les segments distaux.
Les aesthètes sont également de plusieurs types morphologiques, en forme de massue ou lamelle étroite. Elles sont supposées être des chémo-récepteurs (Cf. in Mauchline, 1998, p.18).
Les soies et aesthètes qui occupent des sites particuliers sur les segments et un aspect séquentiel déterminé a permis à Fleminger (1985) de définir une chétotaxie chez les Calanidae.
L'une des fonctions de l'antennule est la détection de la nourriture, des mouvements de l'eau, des prédateurs (Landry, 1980; Poulet et al., 1986; Yen, 1987; Jonnson et Tiselius, 1990; Yen et al., 1992; Yamazaki et Squires, 1996).
Une autre fonction est liée à la reproduction principalement chez le mâle: détection chimique de phéromones et/ou mécanique de la femelle (Fleminger, 1967), puis capture de la partie postérieure de la femelle.
2 - Dans le groupe des Cyclopoida le nombre des segments chez la femelle est très variable, généralement compris entre 6 à 21, parfois supérieure (26 chez Cyclopicina). La longueur de l'antennule atteint rarement la fin du corps, voire du prosome chez les espèces planctoniques marines comme Oithona, mais la fusion (ou perte) de segments confèrent à ceux-ci un apparent allongement, et sont munis de longues soies. Chez les espèces dulçaquicoles ou saumâtres, dont la nage est le plus souvent intermittente, en repos sur des substrats, elles ne dépassent guère le céphalosome, et leurs soies sont relativement courtes.
Chez le mâle, les segments, certains dilatés, montrent plus de fusions que chez la femelle, le nombre maximal étant de 17 chez Macrocyclops albidus. Elles sont géniculées des deux côtés formant vers la partie distale une pince plus ou moins nette, dont la fonction est, comme chez les Calanoida, le maintien de la femelle lors de l'accouplement.
Notons que dans la famille des Thespesioppsyllidae, incluse jusqu'à Huys et Boxshall (1991) parmi les Monstrilloida, les antennules femelles montrent respectivement chez les femelles et les mâles: 15 et 14 segments.
3 - Dans le groupe des Harpacticoida le nombre des segments est variable, de 3 à 9 au maximum chez la femelle et 14 chez le mâle.
La longueur est toujours réduite, dépassant rarement le céphalosome en relation avec le type d'habitat benthique ou constitutif de la meiofaune.
Les antennules du mâle, géniculées des deux côtés, présentent une complexification croissante: de type haplocère pour les moins modifiées (comme Microsetella norvegica), intermédiaire du type sub-chélicère (comme chez Eucanuella spinifera), enfin le type chélicère avec le segment antérieur à la géniculation épais et fortement dilaté et un seul segment postérieur à la géniculation (comme Tigriopus brevicornis)
4 - Dans le groupe des Poecilostomatoida (principalement commensal et parasite) le nombre de segments est réduit (de 6 à 8), légèrement dimorphique sexuellement, mais ne présente pas de géniculation chez le mâle dont elle ne joue aucun rôle lors de l'accouplement.
Bien que les espèces libres soient pélagiques, elles sont très courtes et munies de peu de soies.
5 - Dans le groupe des Siphonostomatoida (essentiellement parasite), le nombre maximal de segments est de 21, souvent moins. Chez le mâle, elles peuvent présenter ou non une géniculation.
6 -Dans le groupe des Monstrilloida, les segments sont souvent mal définis, au nombre de 4 chez la femelle et 5 chez le mâle, chez lequel existe une géniculation bien marquée. Les soies sont variées. Les antennules sont dirigées vers l'avant, quasi parallèles
7 - Pour l'ordre des Gelyelloida, représenté par des espèces présentes dans les milieux souterrains des systèmes karstiques voir in Huys et Boxshall, 1991 (p.185-191)
|



 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :