


Introduction | Morphologie générale | Morphologie des diverses parties du corps et des appendices | Dimensions | Formes | Principales modifications structurelles | Les antennules | Les antennes | Le cadre buccal : Labrum et labium | Les Mandibules | Les Maxillules ou premières maxilles | Les Maxilles (Maxilla) ou secondes maxilles | Les Maxillipèdes | Les pattes natatoires | Variation du nombre des segments corporels | Nomenclature des parties du corps et Métamérie | Structures et organes tégumentaires | Organes visuels | Variations de la morphologie | Sex-ratio | Gynandromorphisme et intersexualité | Accouplement | Parthénogénèse | Caractéristique de la ponte | Les formes larvaires | La durée du développement | La croissance | Régénération | Anomalies | Classification et Phylogénie | Références | Photothèque
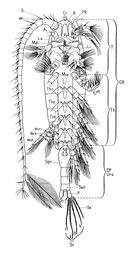

Durant les stades naupliens les copépodes présentent une seconde antenne biramée qui leur sert d'appendice locomoteur (comme les antennules et mandibules). L'exopodite comprend un plus grand nombre de segments que l'endopodite légèrement plus court. Ce caractère primitif demeure à l'état adulte chez des copépodes libres comme Calanus finmarchicus où il semble avoir conservé sa fonction locomotrice , au moins partiellement (fig. 1 - Chez les Calanoida, les antennes sont toujours biramées, l'endopodite et l'exopodite de longueurs plus ou moins égales et plurisegmentés. L'endopodite est généralement trisegmenté avec une fusion plus ou moins nette du segment distal, mais généralement moins par fusion que seule l'insertion de groupes de soies permettent d'individualiser. L'exopodite est multisegmenté à 7 ou 8, voire 9 segments (Euchirella messinensis) Chez les Calanoida, il existe des espèces qui ont une réduction de la taille de l'exopodite (Acartia) 2 - Chez les Harpacticoida, cette tendance est encore plus accentuée, pouvant conduire à une taille insignifiante. Quand il existe le nombre des segments peut être de 1 à 8 (généralement 4 ou moins). L'endopodite présente 3 segments, le premier pouvant être soudé au basis formant alors un allobasis (Lang, 1948, p.46). L'antenne est relativement plus massif et raccourci, moins soyeux que chez les Calanoida, munie d'épines et de denticulations, en relation avec le mode de vie libre mais benthique du groupe. Chez les formes quelque peu nageuses, on observe que l'exopodite est encore bien développé.
3 - Chez les Cyclopoida libres (fig.
4 - Chez les Poecilostomatoida libres, le coxa et le basis sont fusionnés en un coxobasis avec une soie distale, suivi de 2 ou 3 segments, le distal armé de une ou plusieurs soies en crochets. L'exopodite est absent.
5 - Chez les Siphonostomatoida libres, le coxa et le basis, sans soies, peuvent être fusionnés, de même que le premier segment de l'endopodite, et présenter (Hyalopontius, Pontoeciella) ou non (Ratania) un exopodite réduit à un petit segment. L'endopodite bien développé à 2 segments, le distal présentant une forte soie crochue (Hyalopontius), plus plusieurs petites soies à longs crochets terminaux (Ratania). 6 - Dans l'ordre des Monstrilloida les antennes ont totalement disparu comme les autres appendices buccaux chez l'adulte.
|

|
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 16 avril 2026] © copyright 2005-2026 Sorbonne Université, CNRS |
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :Mentions légales et informations nominatives
