Introduction | Morphologie générale | Morphologie des diverses parties du corps et des appendices | Dimensions | Formes | Principales modifications structurelles | Les antennules | Les antennes |
Le cadre buccal : Labrum et labium | Les Mandibules | Les Maxillules ou premières maxilles | Les Maxilles (Maxilla) ou secondes maxilles | Les Maxillipèdes | Les pattes natatoires | Variation du nombre des segments corporels | Nomenclature des parties du corps et Métamérie | Structures et organes tégumentaires | Organes visuels |
Variations de la morphologie | Sex-ratio | Gynandromorphisme et intersexualité | Accouplement | Parthénogénèse | Caractéristique de la ponte |
Les formes larvaires | La durée du développement | La croissance | Régénération |
Anomalies | Classification et Phylogénie | Références | Photothèque
 Gynandromorphisme et intersexualité : Gynandromorphisme et intersexualité :
Les cas de gynandromorphisme ne sont pas exceptionnels chez les copépodes. Ils portent le plus souvent sur la segmentation de l'abdomen, la structure de la cinquième paire de pattes et l'antennule.
Conover (1965) décrit un mâle présentant une masse rouge-orangée dans le céphalosome, typique de celle des femelles gravides. À l'opposé Dussart (1967) signale chez Eudiaptomus gracilis (calanoïde des eaux douces) un cas d'hypermasculinisation chez un mâle dont les deux antennules étaient géniculées. Chez cette espèce, certains individus peuvent présenter une morphologie interne femelle, mais un phénotype masculinisé présentant une antenne droite géniculée et une cinquième paire de pattes modifiée, des spermatohores étant fixés sur l'orifice génital.
Chez Acartia tonsa des mâles génétiques ont été notés comme devenant des femelles phénotypiques durant la croissance (Heinle, 1970).
Fleminger (1985) observe sur les antennules des femelles de Calanidae de la même espèce un nombre et une disposition des soies sensorielles (aesthètes) différents. Les femelles qui présentent le phénotype mâle ont une gonade qui se développe en ovaire et sont fécondées comme les autres, elles dériveraient de mâles génotypiques. Le dimorphisme comme le changement possible de sexe font l'objet de diverses hypothèses émises par cet auteur et qui ne peuvent être résumées ici. Chez Euchaeta (Calanoida), Hopkins (1982) observe le contraire pour des stades juvéniles (femelle génotypique -> mâle phénotypique).
Les copépodes parasités par des Blastodiniens, outre la castration parasitaire qui est de règle, montrent souvent des cas plus ou moins marqués de gynandromorphisme chez lesquels les caractères femelles dominent. Pour Sewell (1951) il y aurait chez la femelle castrée production d'une hormone morphogène mâle susceptible d'affecter le développement dans plusieurs directions, et à divers degrés selon le stade auquel se situe l'infestation. En même temps il pourrait y avoir diminution ou absence d'une hormone femelle, si bien que l'évolution normale aboutissant aux caractères sexuels secondaires ne peut se poursuivre, l'individu retenant certains aspects juvéniles. Dans le genre Corycaeus (Poecilostomatoida), le cas du genre ou sous-genre Farranula (anciennement Corycella) est démonstratif à cet égard, puisqu'il pourrait être considéré comme dérivant du premier par néoténie. les diverses espèces de Farranula ne seraient que des variants modifiés à des degrés divers de la seule espèce F. rostrata.
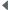 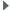 |
|



 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :