Introduction | Morphologie générale | Morphologie des diverses parties du corps et des appendices | Dimensions | Formes | Principales modifications structurelles | Les antennules | Les antennes |
Le cadre buccal : Labrum et labium | Les Mandibules | Les Maxillules ou premières maxilles | Les Maxilles (Maxilla) ou secondes maxilles | Les Maxillipèdes | Les pattes natatoires | Variation du nombre des segments corporels | Nomenclature des parties du corps et Métamérie | Structures et organes tégumentaires | Organes visuels |
Variations de la morphologie | Sex-ratio | Gynandromorphisme et intersexualité | Accouplement | Parthénogénèse | Caractéristique de la ponte |
Les formes larvaires | La durée du développement | La croissance | Régénération |
Anomalies | Classification et Phylogénie | Références | Photothèque
 Les pattes natatoires : Les pattes natatoires :
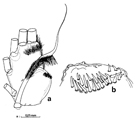
Leur nombre et leur forme vont dépendre du mode de fusion de certains somites, ainsi que des caractéristiques sexuelles.
Le nombre est fonction de la tagmose de la partie génitale qui affecte les segments thoraciques postérieurs.
Les appendices thoraciques sont fondamentalement des appendices natatoires, mais, comme on l'a vu précédemment, la céphalisation du premier somite thoracique a entraîné une modification de la première paire en maxillipède. La seconde paire pouvant aussi être affecté, bien qu'à un moindre degré. Il en est de même pour les deux paires d'appendices thoraciques postérieurs qui sont très variables selon les groupes et le sexe.
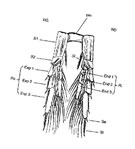 La structure, selon le schéma de Hansen (in Gurney, 1931), est réductible à l'appendice biramé primitif. Il comprend trois segments basilaires, le premier ou Précoxa étant peu identifiable, réduit à un sclérite à la base et latéralement au second segment. Celui-ci ou Coxa (anciennement désigné comme basipodite 1) possède, comme les segments suivants, une soie interne et une épine ou soie externe, ce qui apparaît, compte tenu des fusions ou modifications des segments, un caractère archaïque qui peut être utilisé comme un indice de l'évolution des formes. Le Basis (anciennement basipodite 2) porte une rame interne (Endopodite) et une externe (Exopodite), chacune à trois segments au plus &n(fig. La structure, selon le schéma de Hansen (in Gurney, 1931), est réductible à l'appendice biramé primitif. Il comprend trois segments basilaires, le premier ou Précoxa étant peu identifiable, réduit à un sclérite à la base et latéralement au second segment. Celui-ci ou Coxa (anciennement désigné comme basipodite 1) possède, comme les segments suivants, une soie interne et une épine ou soie externe, ce qui apparaît, compte tenu des fusions ou modifications des segments, un caractère archaïque qui peut être utilisé comme un indice de l'évolution des formes. Le Basis (anciennement basipodite 2) porte une rame interne (Endopodite) et une externe (Exopodite), chacune à trois segments au plus &n(fig. G4). G4).
Gurney (1931) a tenté d'établir les homologies entre les segments des pattes natatoires des Copépodes et celles des Malacostracés primitifs. Au total, neuf segments peuvent être identifiés si l'on accepte la fusion précoxa et pré-ischium, et le fait que dans le segment distal de l'endopodite et de l'exopodite, les soies internes chez le premier et les épines externes chez le second, délimitent des segments, donc définissent quatre segments fusionnés.
Les pattes de chaque paire, du moins les quatre premières (maxillipèdes exclus) sont reliées par les bords latéraux internes de leurs coxae, par un fort sclérite, formant une plaque intercoxale, qui impose un battement synchrone des pattes droite et gauche. Cette plaque basale constitue une structure apparemment unique (Manton, 1977) qui a donné à ce groupe de Crustacés son nom de Copépodes (du grec Kopion: petite rame et podos: pied).
Du fait de leur fonction, les différentes paires de pattes natatoires présentent chez un même individu de grandes similitudes, mais aucune, par la fusion de certains segments et leurs sétations, n'est exactement identique aux précédentes et aux suivantes. Cette caractéristique est utilisée dans les faunes comme l'un des critères de détermination de l'espèce.
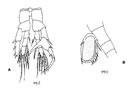
La très grande majorité des formes libres n'ont que cinq paires de pattes, la cinquième, jouxtant le tagme génital, est dans de nombreux cas, non natatoire, profondément modifiée, soit par réduction, pouvant même conduire à leur totale disparition, soit au contraire hypertrophiée, asymétrique, chez les seuls mâles (fig. G12, A).
 Ce changement de fonction, avec pour corollaire une modification structurelle, se manifeste chez le mâle (tout à fait exceptionnellement chez la femelle d'un Harpacticoïde: Phyllopodopsyllus) (fig. G11, B), introduisant un dimorphisme particulier de l'appendice ( utilisé fréquemment dans les faunes). Ceci a lieu lorsque la cinquième paire de patte (maxillipède exclu du numérotage) du mâle est adaptée pour intervenir dans les activités reproductrices: préhension de la femelle, détection de l'aire génitale de celle-ci, manipulation du spermatophore. Ce changement de fonction, avec pour corollaire une modification structurelle, se manifeste chez le mâle (tout à fait exceptionnellement chez la femelle d'un Harpacticoïde: Phyllopodopsyllus) (fig. G11, B), introduisant un dimorphisme particulier de l'appendice ( utilisé fréquemment dans les faunes). Ceci a lieu lorsque la cinquième paire de patte (maxillipède exclu du numérotage) du mâle est adaptée pour intervenir dans les activités reproductrices: préhension de la femelle, détection de l'aire génitale de celle-ci, manipulation du spermatophore.
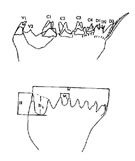
Chez certains Copépodes, et très généralement dans les groupes des Harpacticoïdes et Cyclopoïdes, la cinquième paire est très réduite et il est difficile d'en interpréter les parties. Elle est constituée d'une partie basale courte, en lame ou subcylindrique, portant une ou plusieurs soies ou épines. De part et d'autre de l'orifice des oviductes, on observe parfois une sixième paire de pattes encore plus réduite que la précédente, formant deux saillies, de part et d'autre de l'axe médian, portant des soies.
La perte de fonction de cette sixième paire de pattes aurait permis l'utilisation des muscles pour l'opercule génital, lui-même reliquat d'une paire de pattes qui ferme les ouvertures génitales (Huys et Boxshall, 1991). Des pattes natatoires normalement construites étaient sans doute incompatibles avec l'attache du ou des ovisacs sur le tagme génital, et avec la présence du spermatophore.
Vers l'avant, la première paire de pattes natatoires (P1 dans la nomenclature conventionnelle) montre des variations qui peuvent porter sur une légère diminution de la taille par rapport aux suivantes et une réduction de l'endopodite. Par ailleurs cette première paire de pattes peut subir des changements de structure qui la font ressembler aux maxillipèdes.
Chez les Harpacticoida, Lang (1948, p.64) fournit un tableau de figures comparatives significatives de ces variations.
Des changements similaires se produisent, bien que plus rarement, sur la deuxième paire de pattes (P2) par exemple chez l'Harpacticoïde Canthocamptus. Pour Kabata (1979) ces modifications ont lieu chez les formes qui présentent déjà des appendices préhensiles (A2, Mxp) et servent à accroître les capacités d'accrochage.
L'étude du comportement locomoteur en fonction de l'environnement, souvent mal connu, est pourtant indispensable pour la compréhension de la variation des structures.
  |
|



 Any use of this site for a publication will be mentioned with the following reference :
Any use of this site for a publication will be mentioned with the following reference :